|
|
The propagated action potential along a nerve is primarily dependent on the different concentrations of sodium and potassium ions on the two sides of the semipermeable membrane covering the nerve fiber. According to the membrane theory, the "pores" of this surface membrane are permeable to potassium and impermeable to sodium when resting. The resting (polarized) state, in which the inside of the membrane is negatively charged and the outside of the membrane is positively charged, is the result of electrochemical processes in the neurons and serves as a potential source of energy.Chapter 2:
General Principles Of Clinical Neurology
From R. C. Schafer, DC, PhD, FICC's best-selling book:
“Basic Principles of Chiropractic Neuroscience”
Second Edition
The following materials are provided as a service to our profession. There is no charge for individuals to copy and file these materials. However, they cannot be sold or used in any group or commercial venture without written permission from ACAPress.
All of Dr. Schafer's books are now available on CDs, with all proceeds being donated
to chiropractic research. Please review the complete list of available books.Overview The Evolution of Neurologic Theory Scientific Theories and Principles of Chiropractic Basic Principles Clinical Efficiency Neurologic Aspects of Homeostasis Neurologic Aspects of Immunogenesis Pain Control by Spinal Manipulation Basic Neuroembryology The Neural Tube Somites Basic Neurocytology The Neuron Neuroglia Connective Tissue Electric Changes in Axons Axoplasmic Flow Interneuron Communication Gross Neurologic Areas and Levels Basic Considerations The Brain as a Whole The Skull The Meninges The Ventricular System The Major Cranial Blood Vessels The Supratentorial Level Cranial Nerves I and II The Posterior Fossa Level The Longitudinal Systems The Spinal Levels The Peripheral Levels Microneuropathologic Reactions: General Considerations Basics Cerebral Edema Neuronal Pathology Oligodendroglia and Schwann Cell Pathology Microglia Pathology Astrocyte Proliferation Neuronal Electroneurophysiology Basic Considerations Resting Potential Action Potentials Synaptic Transmission Clinical Applications Importance of Normal Findings Applied Neuroembryology Pathoanatomical Correlations Transient Physiologic Disorders Neural Regeneration BibliographyChapter 2: General Principles of Clinical Neurology
The nervous system and the endocrine system work as partners to provide the majority of functional control for body processes. Guyton, the renowned physiologist, describes the basic function of the nervous system to be the controlling factor for rapid activities such are necessary for muscle contraction, rapidly changing visceral events, and the rate of endocrine secretions.
The dominant action of the nervous system over the physical processes of the body is called neurarchy. In contrast to the nervous system, the endocrine system principally regulates the metabolic functions of the body and controls prolonged physiologic activities.
OVERVIEWThe demanding role of the nervous system of the human body can be appreciated by recognizing that during every minute of life the nervous system must receive thousands of signals from a countless variety of sensory organs, integrate the data, prepare necessary responses, and effect the responses via a multitude of motor and/or autonomic efferent mechanisms. Thus, a specialized network of nerve tissue permeates the body in such a manner that some parts receive and respond to stimuli from the external or internal environments, some parts transmit signals to and from integrating and coordinating centers, and some parts conduct messages from centers peripherally to muscles, vessels, or glands to effect an action.
As an aid in understanding the structural roles involved, physiologists McNaught/Callander portray the general design of the body as consisting of synchronized master tissues and vegetative systems. The quick-acting master tissues specialize in receiving messages from the external and internal environments and reacting to them (eg, nerve and muscle tissues).
Specialized peripheral receptors such as the telereceptors and contact receptors are impressed by stimuli from the external environment, while deep proprioceptors in muscles and joints and the interoceptors and chemoreceptors of the viscera are impressed by stimuli arising within the internal environment.
The slower-acting vegetative systems (eg, digestive, respiratory, circulatory, excretory systems) provide the basic utilities of life necessary for cellular nutrition, growth, or repair and the removal of waste products. These systems are largely regulated by the sympathetic and parasympathetic divisions of the autonomic nervous system, whose activities are modified, balanced, and integrated by centers within the central nervous system (CNS) to meet the constantly changing needs of the body.
The Evolution of Neurologic Theory
The early roots of neurology in the history of health science are extremely cloudy, but bits and pieces have been found by historians and reported by Singer, Majno, and others.
Early Foundations
During the 4th century BC, Plato believed that health disturbances were the effect of disturbed basic elements within the body, which he labeled air, earth, fire, and water. Aristotle taught that the center of sensations of the body was within the heart. He believed that the heart received ripples from the periphery and referred them through blood vessels.
Although the School of Hippocrates later placed the center of consciousness in the brain, the subject remained controversial until the 3rd century BC when teachers at the learning center of Alexandria distinguished nerves from tendons and blood vessels, and some motor and sensory functions within peripheral nerves were established.
The early Greeks taught that nerve functions were accomplished by the movement of "humors" within the body, and this belief prevailed until the 17th century in most Western countries. Claudium Galen, in the 2nd century AD, accepted the general principles established at Alexandria and added knowledge about the spinal cord that was so detailed it remained unchanged until the 19th century. Galen was also the first to teach the proper positions and relationships of the vertebrae to the spinal column and the significance of the pulse and the arteries.
During the 19th century, physiologists began applying the new knowledge of electricity to neural function and muscle contraction, and action potentials were established in nerves and muscles in the 1800s. Most of the data collected at this time were manifestations following severed nerves. Schwann described nerve fibers and cells in 1839, but the segmental structure of nerve fibers and demyelination processes were not defined by Ranvier until the 1870s.
The all-or-nothing principle of neuromuscular excitability was not firmly established until 1871 by Bowditch for heart muscle, until 1909 by Lucas for skeletal muscle, and until 1922 by Adrian/Forbes for nerves. It should be pointed out that D. D. Palmer established the basic principles of chiropractic in the 1890s, in the middle of these rudimentary findings.
Contemporary Developments
With the development of the vacuum tube in the 1920s, neurologic data expanded rapidly because measurements could be made in millisecond and microvolt ranges. The modern views of the relationship between neural structure and function were established during the period between 1900 and 1920, as were concepts of chemical actions at the junctions between excitable cells, but the compound action potential of nerve tissue was not proved until 1937. The ionic basis of the action potential in single nerve fibers was established in 1952. With the development of the electron microscope during the 1950s, the investigation of intricate neurologic morphology began.
Most of the facts proved during this century evolved from theories established many decades before. In reviewing the history of neuroscience, it is amazing to see how often the speculations of scientists with disciplined imaginations, who had to base their theories primarily on empiric experiences, proved quite accurate once technology advanced to the point where their hypotheses could be proved.
The science of chiropractic is no exception to this pattern. The lack of theoretical explanations has never been a problem. The challenge has been to formulate hypotheses in such a manner that they can be tested and to develop the means to do so under controlled conditions.
Scientific Theories and Principles of Chiropractic
As a science, chiropractic and other licensed health-care professions are based upon facts demonstrated within the basic sciences; and as healing arts, many procedures are based on empiric evidence and scientific theories and principles that await indisputable confirmation.
In discussing the evolution of neurology and the concept of chiropractic, Haldeman/Hammerich show how chiropractic concepts have evolved in parallel with increasing knowledge of neurologic anatomy and physiology. Likewise in osteopathy, Denslow explains that much is known, much is unknown, and much is controversial about the osteopathic lesion (subluxation). The same is true for chemotherapy.
Basic Principles
Most authorities will agree with Vear when he states that the scope of chiropractic practice is inexorably tied to the biologic concept that relates structure to function and how that concept relates to health and disease.
Although not limited to these by far, it can be stated that the core of the chiropractic approach is built upon three related scientific principles that have been published within the ACA booklet Chiropractic State of the Art for many years. They are listed below with slight modification to be current with terminology in use today.
Disease may be caused by disturbances of the nervous system. While many factors impair health, disturbances of the nervous system are among the most important factors of disease etiology. The nervous system coordinates cellular activities for adaptation to external or internal change. Agents and conditions that irritate the nervous system, and to which the body cannot successfully adapt, produce changes in the pattern of nerve impulses deviating from the norm. Thus originate many functional disturbances, which, if left unchecked, can lead to disease processes.
Disturbances of the nervous system may be caused by derangements of musculoskeletal structures. Off-centerings (subluxations) and fixations of vertebral and pelvic segments represent common mechanical clinical findings. Extended abnormal involvement of the nervous system may result from disturbances, strains, and stresses arising within the musculoskeletal system due to an individual's attempt to maintain various static and dynamic postures. The mechanical lesion (subluxation) is a common result of gravitational strains, asymmetrical activities and efforts, developmental defects, or other mechanical, chemical, or psychic irritations of the nervous system.
Disturbances of the nervous system may cause or aggravate disease processes in various parts or functions of the body. Vertebral and pelvic subluxations may be involved in common functional disorders of a visceral and vasomotor nature and at times may produce phenomena that relate to the special organs. Under predisposing circumstances, almost any component of the nervous system may directly or indirectly cause reactions within any other component by means of reflex mediation. The conjunction of independent causes of bodily dysfunction may jointly have more serious debilitating effects than a singular cause might have separately. A subluxation complex may contribute to the "triggering" or exacerbating of certain types of neurovascular and neurovisceral instabilities, and correction of the lesion(s) is often an imperative toward effective total management of the case.
Clinical Efficiency
Support of the efficacy of chiropractic theory and practice within the scientific community has grown steadily in recent years. Janse, Biederman, and Weiant, among others, reinforce their discussions on this topic with numerous quotations from American and European medical doctors.
In discussing the scientific basis for chiropractic, Inglis and associates report in their well-documented monograph that chiropractic theory has been confirmed to be sometimes more effective than medical treatment in many cases. Brunarski comes to the same conclusion after reviewing a large number of clinical trials, and points out that bias against manipulation may have missed important differences in treatment effectiveness. More recently, Rupert and associates describe the results of chiropractic adjustments in a controlled clinical trial conducted in Egypt. Many similar studies are being planned today.
Wayne has documented the effects of a subluxation on the functional efficiency of the body, and Haldeman has emphasized the importance of nerve pressure, afferent input from paraspinal structure, protective reflexes, and the somatovisceral reflex. Similar concepts are reported by others.
In writing of the theory of manipulative medicine, Pinkenburg stated that current evidence strongly suggests that sensory afferent impulses may play an important role in the maintenance of dynamic equilibrium of the human body in its environment. Overstress of muscles, tendons, and ligaments causes a change in patterns of afferent nerve impulses that are fired from their nociceptors and mechanoceptors and transmitted to the central nervous system (CNS). It can be hypothesized that, as a result of these changes, stress develops that affects not only the body's dynamic equilibrium with the forces of gravity but also possible visceral dysfunction, and, in turn, visceral disorders influence the body's dynamic equilibrium.
In reviewing musculoskeletal factors in chronic disease, Thorpe points out that many chronic diseases can be helped by manipulative therapy. He concludes that because the spinal segments are extremely sensitive, and since they are interrelated with the viscera and muscles, they affect body reflexes in cases where chronic disease exists.
The efficiency of chiropractic care in mental/emotional disturbances has also been documented. The most notable of these are from Quigley, regarding patients at the Clear View Sanitarium; the many writings of H. S. Schwartz, especially his analysis of 350 mental patients treated by chiropractors; and Mears' paper on cervical spine distortion. Guze, Homewood, Kimmel, and Sulzer have also reported favorably on the role of chiropractic in structural psychotherapy and somatopsychic disorders.
Neurologic Aspects of Homeostasis
Homeostasis is defined in Taber's Cyclopedic Medical Dictionary as "the state of equilibrium of the internal environment of the body that is maintained by dynamic processes of feedback and regulation." Stedman's Medical Dictionary defines homeostasis as "the state of equilibrium (balance between opposing pressures) in the body regarding various functions and to the chemical composition of the fluids and tissues." Dorland's Medical Dictionary simply defines it as "a tendency to stability in the normal physiologic states of the organism." Hoag explains that the term homeostasis should not suggest a fixed state of physiologic balance, but rather, a state of dynamic equilibrium. He also explains how the musculoskeletal system is important in its maintenance.
The restoration of homeostasis was written about extensively in early chiropractic literature, and it still is today, especially by those who emphasize systemic (primarily autonomic) balancing. Frequent examples are seen within the current literature of those who apply acupuncture, applied kinesiology, or the principles of Bennett's neurovascular reflexes.
The term is infrequently used in medical literature except by physiologists. For example, Guyton refers to the process throughout his text whenever he describes a functional control system or mechanism. Pottenger felt that the diencephalon was the central control center for homeostasis, as it seemed to be a major receiving and distributing center for nerve impulses that arise in any part of the body.
The necessity of homeostasis to maintain health is often referred to in the writings of chiropractic educators Harper, Janse, Vernor, and D. D. Palmer, but the subject is rarely indexed. It can be assumed that the many references to innate intelligence within pioneer chiropractic literature were reflections of homeostatic processes. Homewood also makes this assumption. Jacquet refers to homeostasis in terms of bioenergetics and explains what Lowen calls "the inner language of the body."
Strang is one of many current chiropractic educators who states without hesitation that "Chiropractic rationale begins with the law of biology known as homeostasis," and he offers considerable text to relate its importance to diagnosis and case management within chiropractic. He explains that:(1) homeostasis enables the body to stay in balance in an ever-changing environment,
(2) the nervous system is the prime controller of homeostasis, and
(3) faulty musculoskeletal relationships can cause dysfunction in the nervous system.In addition, he quotes the following that is attributed to Janse: "Normal integrated neurological conduct equals homeostasis and health, and disturbed neurological conduct results in pathophysiology, disintegration of homeostasis, and eventually the intrusion of disease."
Neurologic Aspects of Immunogenesis
It has been strongly felt by most chiropractic practitioners that periodic chiropractic adjustments, even when the patient is in a healthy state, will do much to enhance the patient's resistance to adverse forces. While allopathy has placed great emphasis on the ability of environmental pathogens to attack a host, chiropractic's approach has been directed to the ability of a host to ward off most such attacks if a healthy homeostasis exists. Obviously, both viewpoints are worthy of consideration being opposite sides of the same coin.
Some microorganisms are highly virulent and severely attack even the most apparently healthy individual. The AIDS virus is a current example. On the other hand, we do not live within a sterile environment. We are constantly exposed to large quantities of potentially dangerous bacteria, viruses, fungi, chemicals, etc, within the air we inhale and the food we ingest without noticeable effect. Streptococci can be shown in the nasopharynx of almost everyone, yet it is obvious that few people manifest clinical "strep throat."
The conclusion can easily be drawn that assisting the patient's vis medicatrix naturae has always been the objective of mainstream chiropractic. Evidence to confirm the efficiency of chiropractic care in this area of preventive health is essentially empiric so far, yet it is based upon some rational scientific principles.
Several years ago, Zhigalina pointed out that the vagus nerve is considered the efferent path for the effect of the CNS on immunologic reactions. Stimulation of the vagus produces an increase in the titer of antibodies, a rise in leukocytes, an increase in phagocytic activity, a decrease in serum protein, and a redistribution of its fraction. He also explains how vagal stimulation is capable of rising the immunologic reaction of the organism and may be used in applied immunology. In this text, Gordienko and associates state that antigens can excite peripheral receptors, to cause transformation of these excitations into propagated impulses, and to cause well-defined changes of the biopotentials in the sensory nerve.
It was also found that different oscillation of potentials was obtained with different antigens, and it appeared that certain antigens in contact with skin receptors choose their excitation, which is transmitted by the nerves and may play a decisive role in the organization of the defensive processes of the organism against the initial infection.
Although many of Gordienko's experiments added great credibility to chiropractic hypotheses, one in particular should be described: A rabbit's ear was prepared by isolating the blood circulation but leaving the innervation intact. An antigen was introduced intracutaneously, and this lead to the production of specific agglutins when the antigen contacted the skin receptors for 1/2--1 minute. Because the blood circulation had been restricted, these results attest to the possibility of the reflex production of specific agglutinins on the intracutaneous introduction of an antigen.
Gondienko's group also showed that skin receptors of the immunized animals react to specific antigens for a longer duration and often more powerfully. The change in reaction proved again that immunogenesis is ensured above all by stimulation of the nervous system and initiation of its reflex mechanisms.
It has long been held empirically within chiropractic that even subclinical states of malnutrition have an adverse effect upon the nervous system. In this context, the relationship between nutrition and infection established by Faulk and associates, among others, is significant. They determined that nutrition and infection interact in two ways:(1) malnourished people are more prone to infection and
(2) infection may predispose malnutrition.Pain Control by Spinal Manipulation
As most patients enter a chiropractor's office with a complaint of pain, the relief of pain has always been a common goal in chiropractic therapeutics. The excellent results achieved have generally been attributed to the reduction of nerve irritation, muscle spasm, edema, and circulatory stasis. However, Vernon and associates have recently expanded our view by showing, under controlled conditions, that plasma levels of endorphins (intrinsic opiate-like compounds) are increased after chiropractic spinal adjustments. These and other effects of chiropractic adjustments will be described in following chapters.
BASIC NEUROEMBRYOLOGYA brief review of neural embryology is helpful to appreciate that the locations of structures in the adult reflect an orderly development from their primitive states.
The development of the nervous system is a gradual process of cell division, migration, remodeling, and specialization, which results in neural circuits requiring cell-to-cell recognition. The mechanisms responsible for this cell-to-cell recognition remain relatively unknown as compared to the sequence of events that occur.
The CNS at birth has completed most of its early stages of cell division, migration, and specialization. Most of the neural circuitry has been laid down. Neuroblasts are continuing to divide only in limited brain regions, according to Bishop. Most mammals, therefore, have a nearly full complement of neurons at birth. Nonetheless, the functional capabilities of the CNS of the newborn have little resemblance to those of the adult. Postnatal maturation must proceed in the proper sequence and at the proper rate if CNS development deficits in the adult are to be avoided.
The Neural Tube
During the 3rd week of development, the embryonic disk consists of two rapidly differentiating layers and a thickened area (primitive streak) from which the notochord is an extension. Ectoderm covering the folds of the notochord soon fold inward to form the lateral grooves of the primitive neural tube. It is from this basic structure that the entire CNS develops. The cells lining the neural tube become the nerve tissue of the brain and spinal cord. The canal itself becomes distended with fluid to form the ventricles of the brain and central canal of the spinal cord.
Longitudinal Differentiation
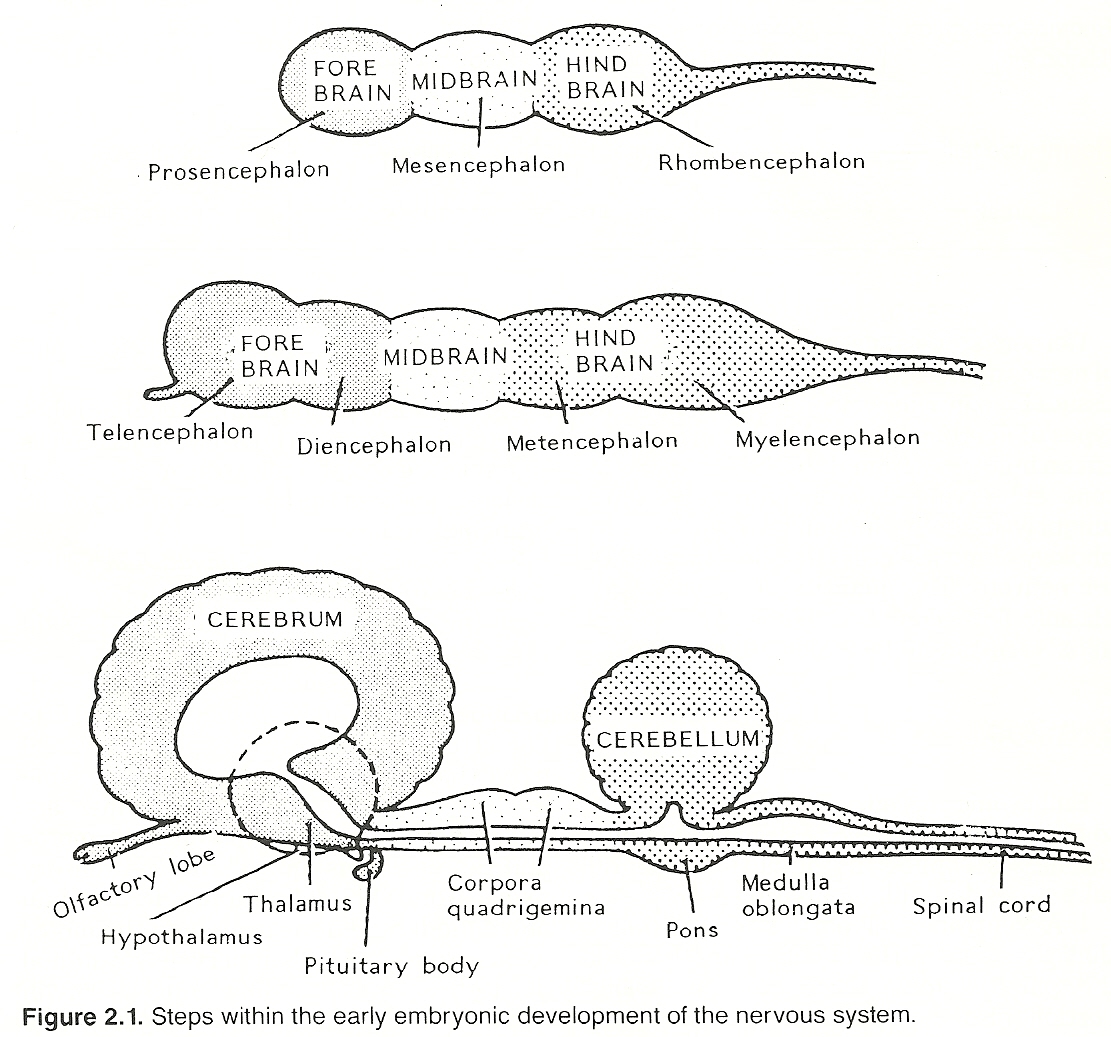
The ectodermal neural tube is formed between the 18th and 25th day of gestation. During the first stage of differentiation, the distended body of the neural tube begins to constrict to form three primary vesicles: the cephalad forebrain (prosencephalon), midbrain (mesencephalon), and caudad hindbrain (rhombencephalon).
During the second stage, the forepart of the tube constricts and becomes the telencephalon (cerebral hemisphere area) and diencephalon (thalamus and hypothalamus area), and the hindpart constricts and becomes the metencephalon (midbrain and cerebellum area) and myelencephalon (medulla area). Even before the neural tube closes, the tail of the body of the neural tube begins to develop into the spinal cord. These subdivisions are the precursors of the four major anatomical levels in the adult. See Table 2.1.
Transverse Differentiation
During the stages of longitudinal differentiation, transverse differentiation also occurs. The alar plate develops on the dorsal aspect (sensory functions), mesodermal tissues grow into the neural tube and form blood vessels, and the basal plate forms on the ventral aspect (motor functions). These transverse divisions later become the major longitudinal functional systems in the adult (ie, the sensory, motor, visceral, vascular, consciousness, and cerebrospinal fluid systems).
Peripheral Differentiation
Cells continue to divide and split away from the neural tube as it closes to form two columns (neural crests) along the junction between the surface ectoderm and the neural tube. Cells within these crests multiply, migrate laterally, and differentiate to form the dorsal root ganglia (cell bodies of sensory neurons), autonomic ganglia (cell bodies of visceral neurons), Schwann cells for myelination and its neurilemma, and outgrowths of the neural tube (axons and dendrites). The neural crest also gives rise to the adrenal medulla.
Table 2.1. Progression of Major Structures from the Neural Tube___ ___ | Rhinencephalon | | | Basal ganglia | | Telencephalon —> | Cerebral cortex | | | Lateral ventricles | Prosencephalon | --- | (Forebrain) | | Epithalamus | Supratentorial | | Thalamus | level | | Hypothalamus | | Diencephalon —> | Optic nerves | | | Neurohypophysis | --- | Pineal gland | | Third ventricle | --- ___ ___ | Mesencephalon | Mesencephalon | Midbrain | (Midbrain) | | | --- ___ | Posterior fossa ___ | Cerebellum | level Rhombencephalon | Metencephalon | Pons | (Hindbrain) | Myelencephalon | Medulla oblongata | | | Fourth ventricle | --- --- Neural tube tail ———— > Spinal cord and central canal | Spinal level Neural crest ——————> Peripheral nerves and ganglia | Peripheral levelAs development progresses, the cells within the dorsal root ganglia send axons peripherally throughout the embryo and centrally into the alar plate as a CNS connection. Connections between peripheral structures and the CNS are also made by outgrowths of axons from neurons in the basal plate. The cell bodies within the autonomic ganglia project motor axons to and receive sensory axons from the viscera.
Werner's studies suggest that the influence of nerves upon the morphology of peripheral tissues is an intrinsic mechanism of different types of neurons rather than being dependent on influences from the CNS. Sensory neurons subserving different functions have uniquely different influences on the morphology of muscle cells than do gamma motor neurons. The quantity of varying axoplasmic flow along the axons to their terminals appears to be a regulating factor.
Somites
Embryonic mesoderm that lies adjacent to the neural tube becomes segmented into cell masses (somites) as the end of the neural tube closes, later differentiating into:
A ventromedial portion (sclerotome) that gives rise to the cartilage and bone of the spinal column and base of the skull. This portion also incorporates the notochord, whose remnant becomes the nucleus pulposis of the intervertebral discs.
A dorsomedial portion (myotome) that differentiates into primordial muscle cells that migrate peripherally to form all striated skeletal muscle of the body except that of the branchial arches.
A lateral portion (dermatome) that gives rise to cells that migrate peripherally to form the connective tissue layer of the skin.
The largest and fastest fibers in most peripheral nerves are those that bring messages to the CNS about skeletal muscle tension and position. Motor neurons to skeletal muscle are almost as large. The prominence of these types of fibers reflects the adaptive utility of rapid movement. Sensory data from cutaneous structures and viscera travel centrally much slower. Autonomic efferent fibers to smooth muscles and glands are even smaller and slower compared to those of somatic neurons.(1) the normal period,
(2) an absolute refractory period, and
(3) a relative refractory period.
The absolute refractory period. This is a short period following stimulation during which the nerve fiber is not excitable, making it impossible for a fiber to respond continuously to stimuli. Thus, there is a rhythmic discharge of nervous waves (impulses) from receptor to nerve fiber rather than a continuous flow. Small nerve fibers have longer refractory periods than do large fibers.
The relative refractory period. The intensity of a stimulus at the receptor end of a fiber must reach threshold value before it can excite the neuron. During the relative refractory period, the stimulus must be stronger than during the normal resting period. The smaller the fiber, the higher the threshold value of the stimulus must be to produce excitation.Impulse Frequency, Speed, and Strength. The frequency of impulses usually varies directly with the strength of the stimulus. Velocity of conduction, as described previously, varies with the diameter of the neuron's axis cylinder; ie, thicker fibers conduct more rapidly than finer fibers. Although the strength of the electric charge of a neuron varies according to its size, all neurons are governed by the all-or-none law, which states that fibers give a maximum response to a stimulus or no response at all; ie, a stimulus above threshold value will not increase the strength of a single fiber's electric charge.
Chemical Synaptic Inhibition and Extrication
As synaptic function is based on a carefully controlled integration of chemical influences, a slight chemical imbalance (eg, from certain medications, nutritional and electrolytic imbalances) may produce a distinct effect upon synaptic transmission in terms of noxious systemic and behavioral effects. Impulse transmission among billions of neurons, many of which have thousands of transmitter-releasing terminals, is one reason the nervous system is the most complex tissue in the animal kingdom.
Action potential transmission at the synapse is not in the all-or-none fashion. It may be inhibited or amplified according to the specific type of transmitter substance produced. Inhibitory factors can also be produced presynaptically in the axon or produced centrally by hypoxia or drugs. Synapses are more readily fatigued than are nerve fibers, and they are more readily affected by anesthetics and other drugs (eg, nicotine).
When stimulated, some fibers (cholinergic) liberate an acetylcholine substance; other fibers (adrenergic) liberate epinephrine. These neurotransmitters are the two major substances liberated at synapses, but more have been isolated and many more are suspected.
The cholinergic fibers include all preganglionic fibers, all postganglionic parasympathetic fibers, postganglionic sympathetic fibers to the sweat glands, and efferent fibers to skeletal muscle. The adrenergic fibers include nearly all postganglionic sympathetic fibers except those supplying the sweat glands.
A hypersensitive synaptic response to stimuli often follows the depression of higher inhibition centers, where even slight stimulation may be interpreted as pain and mild sounds are perceived as loud noises. While in this condition systemically, an individual is highly susceptible to suggestion beneficial or detrimental.
Electric Transmission
It has recently been shown that some nerves communicate with each other by electric rather than chemical transmission at synapses. The mechanism is probably due to linkage of cytoplasmic bridges between end feet and receptor membranes. The transmission usually has an excitatory effect, but for reasons unknown, it may be inhibitory in some situations.
Unlike chemical transmission, Pappas shows that electric synaptic transmission may be bidirectional; thus, either side may be pre- or post-synaptic at different times. While it has been shown that electric transmission is also faster than chemical transmission, further studies are necessary to determine the exact scope, distribution, and mechanisms involved in humans.
Axoplasmic Flow
During impulse activity, materials are transported from the cell body's endoplasmic reticulum, microtubules, and neurofilaments along the axon via its gel-like axoplasm that is continuously synthesized in the cell body. This axoplasmic flow moves materials slowly up and down the fibers during impulse activity.
Investigations, still in their infancy, have isolated three major types of axoplasmic transport: fast, slow, and retrograde. The exact mechanisms are unknown, but certain facts have been demonstrated. For example, fast transport seems to move polypeptides, proteins, and particulate matter at a rate of 400 mm/day. This rate depends on tissue temperature and oxygen and ATP concentrations; age and fiber function or size are not factors.
It is thought that protein filaments bind to the transported material and creep along the axon by attaching to successive locations on the longitudinally oriented microtubules. If a neuron becomes ischemic, electric excitability and fast transport fail in about 15 minutes.
Slow transport moves at a rate of 1—3mm/day and appears to be linked to the transient binding of mitochondria to the same protein filaments involved in fast transport. Axon mitochondria have been observed to have a constant to-and-from movement with a net distalward drift.
Retrograde transport has most of the properties of fast transport except its rate is about 200 mm/day. It is postulated that it may be the mechanism for the centripetal movement of certain viruses and toxins (eg, rabies, tetanus).
Interneuron Communication
There are four basic ways in which neurons are functionally linked:
One receptor neuron may be linked with one connector neuron that communicates with one effector neuron. This is a rare occurrence.
One receptor neuron may be linked to one connector neuron that relays messages to several effector neurons.
Several receptor neurons may be linked to one connector neuron that communicates with one effector neuron.
Several receptor neurons may be linked to one connector neuron that relays messages to several higher and/or lower effector neurons.
 |
The brain is the largest and most complex mass of nervous tissue in the body. It comprises five connected parts: the cerebral hemispheres, the midbrain, the cerebellum, the pons, and the medulla oblongata.
During embryonic life, the cerebral hemispheres, their commissures, and the 1st, 2nd, and 3rd ventricles are developed from the forebrain. The corpora quadrigemina, the central peduncles, and the cerebral aqueduct are developed from the midbrain. The medulla, pons, cerebellum, and 4th ventricle are developed from the hindbrain.
The weight of the adult brain is about 48.6 oz in the male and about 44 oz in the female. The weight of the brain is an indication of growth, which in early life depends on the enlargement of the cells and their processes, the myelination of certain nerve fibers, and an increase in the amount of neuroglia. The brain grows rapidly up to the 5th year of age and, in general, ceases to grow much beyond the 20th year. In advanced age, the brain gradually loses weight.
The development of the brain is not only a matter of growth but also a matter of forming new pathways; ie, new synapses and a permanent modification of the synapses that are functionally active during various forms of mental activity. The nature of brain tissue and the use to which it is put determine to some extent the length of time during which development may continue.
It appears that mental activity keeps the brain active and capable of development, just as exercising a muscle tends to prevent atrophy or loss of function. This point has been commonly neglected in geriatric care, where the mentally healthy elderly are often treated socially as children.
The Skull
The brain appears firm in illustrations and upon dissection after being injected with a formalin solution. This firmness, however, is not present in vivo. In the living body, the brain is soft but still maintains a certain degree of cohesiveness. Because of its soft consistency, it requires dependable protection against mechanical compression.
Nilsson reminds us that a protective shielding is furnished by the bone-lined cranial cavity. The individual bones forming the cranial vault are the paired temporal and parietal and the single frontal, ethmoid, sphenoid, and occipital bones. Firmly adherent to the external cranial wall, except under its floor, is the scalp. This structure is composed of the skin, the tela subcutanea, the galea aponeurotica, the subaponeurotic soft tissue, and the external periosteum of the bones forming the roof and sides of the cranium.
On the inner cranial wall is the internal periosteum (also referred to by some as the external layer of the cranial dura matter), while next to it is the shallow subdural space containing a small amount of fluid. Then follow the arachnoid meninx and the subarachnoid cerebrospinal fluid. The innermost meninx is the pia mater, which lines the surface of the brain.
Besides the classic brain-protecting structures and elements of the scalp, cranium, meninges, and cerebrospinal fluid, other structures help as shock absorbers to protect the brain. These include joint fat, ligaments, discs, muscles, and the spinal curves that soften or entirely obliterate most sharp jars from harming the brain during body movements.
The Fontanels and Sutures
The anterior and larger fontanel remains about the same size for the first year of life, then diminishes and closes about the 12th month. The posterior fontanel closes in about 6 weeks. In rickets, hydrocephalus, hereditary syphilis, and cretinism, the fontanels and sutures remain open after the normal time for closure.
Normal skull measurements and proportions depend upon the proper sequential closing of the sutures, and most all abnormal head shapes are the result of premature closure. Growth always occurs in a direction perpendicular to the line of the suture, thus inspection and palpation allow us to determine the suture(s) at fault. If one suture closes prematurely, growth occurs in the remaining open sutures in compensation and this results in deformity.
The Meninges
The brain and spinal cord are enclosed within three membranes. They are, from without inward, the dura mater, arachnoid mater, and pia mater.
Dura Mater
The dura mater is a dense membrane of fibrous connective tissue that contains many blood vessels. The cranial and spinal portions of the dura differ, but they form one complete membrane. The cranial dura is arranged in two layers that are closely connected except where they separate to form sinuses for the passage of venous blood (viz, the intracranial venous sinuses). The outer (endosteal) layer adheres to the cranial bones and forms a part of their internal periosteum.
These layers are separated from the bone of the skull by the epidural space. The inner (meningeal) layer covers the brain and sends numerous prolongations inward for the support and protection of the different lobes of the brain. These projections also form sinuses that return the blood from the brain and form sheaths for the nerves that pass from the skull.
In two areas, the dura mater folds into the cranial cavity to form a fibrous barrier:
(1) the falx cerebri, located between the cerebral hemispheres;
(2) the tentorium cerebelli, demarcating the superior limit of the posterior fossa.
Two similar folds of dura should be noted:
(1) the slight falx cerebelli, which attaches to the internal occipital crest and
(2) the diaphragma sellae, which forms a tentorium for the hypophysis cerebri. Beneath the dura mater lies the subdural space.
The spinal dura mater forms a loose sheath around the spinal cord. It consists of an extension of only the inner layer of cranial dura. The outer layer ceases at the foramen magnum, and its role is taken by the periosteal lining of the vertebral canal. Outside the spinal dura is the epidural space, which contains venous plexuses and fat. Between the spinal dura and the arachnoid maters is a potential space (subdural cavity) that contains only enough fluid to moisten their contiguous structures.
Arachnoid Mater
The arachnoid mater is a delicate filamentous serous membrane placed between the dura and pia maters. The cranial portion loosely invests the brain, and, except for the longitudinal fissure, it passes over the various convolutions and sulci and does not dip into them.
The cavity (subarachnoid space) between the arachnoid and the pia maters is occupied by spongy connective tissue and intercommunicating channels. It surrounds the entire brain and spinal cord and is filled with cerebrospinal fluid. Small tufts of arachnoid villi (Pacchionian granulations) invaginate into the dural venous sinuses, especially along the dorsal convexity of the cerebral hemispheres, parallel to the interhemispheric (longitudinal) fissure.
The spinal portion of arachnoid is tubular, loosely surrounds the spinal cord, and contains cerebrospinal fluid.
Pia Mater
The pia mater is the innermost layer of meningeal coverings. It is a vascular membrane, composed of a single cell layer of cells derived from mesoderm, consisting of a plexus of blood vessels held together by fine areolar connective tissue. The cranial portion closely invests the surface of the brain and dips into the convolutions.
The spinal portion of pia mater is thicker and less vascular than the cranial aspect. It is closely adherent to the entire surface of the spinal cord and sends a process into the ventral fissure. To stabilize the spinal cord within the vertebral column, dentate ligaments extend on either side between the origins of the spinal nerves, pierce the arachnoid, and insert into the dura.
While the dural sac and subarachnoid space ends at the S2 level, the pia mater continues downward from the end of the spinal cord as a filamentous membrane (filum terminale) to join the dural sac at the S2 level and then attaches caudally to the posterior surface of the coccyx (sacrococcygeal ligament).
The Ventricular System
The ventricular system consists of the dura, arachnoid, and pia maters; the epidural, subdural, and subarachnoid spaces; and the cerebrospinal fluid (CSF). It exists at the supratentorial, posterior fossa, and spinal levels.
CSF is colorless, alkaline, and has a normal specific gravity of 1.005—1.008. It circulates through the ventricles and the subarachnoid space and consists chiefly of water; a large amount of carbon dioxide; some glucose, salts similar to those found in plasma, and lymphocytes; and traces of protein and pituitary hormones.
Major purposes of CSF are to keep the inner and outer surfaces of the brain and spinal cord moist, to lubricate them, to compensate for fluctuations in cranial blood volume in an attempt to maintain a cranial fluid content constant, to serve as an exchange medium of metabolic substances between nerve cells and CSF, and to serve as a protective hydraulic cushioning and buffering mechanism to disperse forces applied against the cranium or that travel upward in the spine from the lower extremities (eg, during gait, jumps).
The meningeal membranes and spaces filled with CSF form a pad enclosing the brain and spinal cord on all sides. CSF is secreted and diffused from blood by the epithelial cells that surround the choroid plexus of the ventricles. After filling the lateral ventricles, it escapes via the foramen of Monro into the 3rd ventricle and then through the aqueduct into the 4th ventricle.
From the 4th ventricle, CSF passes through the median foramen of Magendie and the two lateral foramina of Luschka, into the subarachnoid spaces, and reaches the cisterna magna. CSF fluid from the cisterna magna also bathes the peripheral parts of the brain in contact with the ventricular system. The subarachnoid space communicates with the interior of the brain via the ventricular system.
CSF fluid also travels along the course of certain cranial nerves, especially the olfactory nerve. This loophole allows the entry of infection from the nasal cavities to the cerebral cavity.
From the subarachnoid spaces, CSF is absorbed into the blood stream through the villi of the arachnoid mater that project into the superior sagittal sinus. A small amount passes into the perineural lymphatics of the cranial and spinal nerves.
The Major Cranial Blood Vessels
The brain is supplied by two pairs of major arteries: anteriorly by the internal carotid and posteriorly by the vertebral. Many of the major arterial channels can be seen on the undersurface of the brain beneath the arachnoid mater.
The Internal Carotid Arteries
The internal carotids ascend the anterolateral neck and pierce the dura mater medial to the anterior clinoid processes, which they groove. They immediately give off
(1) the ophthalamic arteries, which run below the optic nerve through the optic canal and
(2) the posterior communicating arteries, which run posteriorly, medial to cranial nerve III, to join with the posterior cerebral artery.
They then ascend with the optic nerve and end as the anterior and middle cerebral arteries. After running medially above the optic nerve, the anterior cerebral arteries are joined by the anterior communicating artery. Then they continue in the midline between the two hemispheres to supply the medial surfaces of the cerebrum.
The middle cerebral artery runs laterally in the stem of the lateral cerebral sulcus, coursing between the temporal and frontal lobes to emerge upon the surface of the insula (island of Reil), where its branches supply the lateral surface of the hemisphere. A series of anastomotic channels lying at the base of the brain, known as the cerebral arterial circle of Willis, permits communication between the carotid and vertebral systems.
The Vertebrobasilar System
The vertebral arteries ascend through the cervical transverse processes and pierce the dura matter behind the occipital condyle and the margin of the foramen magnum, which they groove. They then project forward to the inferior edge of the pons where they join together to form the basilar artery, whose branches solely supply the caudal structures of the brain (viz, the occipital lobe, cerebellum, undersurface of temporal lobe, thalamus, midbrain, medulla, and portions of the superior spinal cord).
The basilar artery ascends the midline bony slope from the inferior edge of the pons to its upper edge, where it terminates in a T-shaped bifurcation, above cranial nerves III and IV, to form the right and left posterior cerebral arteries. Before this occurs, three pairs of arteries branch horizontally from the basilar artery:
The first and largest branches of the vertebral arteries are the posterior inferior cerebellar arteries. During their course to the cerebellum, they loop around or among cranial nerves IX, X, and XI.
Next, two other smaller paired branches proceed horizontally from the basilar artery:
(1) the anterior inferior cerebellar arteries (that pass above, below, or between cranial nerves VII and VIII), which project labyrinthine branches to the inner ear, and
(2) the superior cerebellar arteries.
Venous Drainage of the Brain
The veins within the skull roughly follow the arterial courses. They meet at enlarged channels (venous sinuses) contained within the two layers of dura mater. The two major sinuses are the superior and inferior sagittal sinuses that run medially between the cerebri in the falx cerebri, merge in the occipital region to form the confluence of sinuses that drains into the transverse and sigmoid sinuses and then exits the skull through the jugular foramina as the internal jugular veins.
The Supratentorial Level
On each side of the cranial vault, the floor of the skull is separated into three fossae (anterior, middle, and posterior). The rigid tentorium cerebelli, which lies nearly horizontal and attaches to the petrous ridges laterally and the occiput posteriorly, divides the posterior fossa from the anterior and middle fossae.
The structures of the nervous system situated above the tentorium cerebelli are those of the supratentorial level, all of which are derivatives of the primitive telencephalon and diencephalon. The major structures at this level are the cerebral hemispheres, the diencephalon, and portions of the olfactory and optic cranial nerves.
The Cerebrum
All lobes of the brain and portions of the brain stem partake in the consciousness system, which is concerned with maintaining wakefulness, awareness, and sleep states. The supratentorial level is also concerned with such faculties as cognition, imagination, intelligence, language, memory, olfaction, perception, vision, and volition. The cerebrum serves are the primary center for memory, motor response patterns, will power, and sensory discrimination.
The two cerebral hemispheres are separated by the longitudinal fissure, and each hemisphere has a convoluted surface. These convolutions have the effect of increasing the amount of cerebral gray matter, which contains a network of intricate layers of interconnecting neuron cell bodies. They number in the thousands of millions, and each cell communicates synaptically with hundreds of others. Depending upon the authority used, 75%—90% of all neuron cell bodies are located in the cerebral cortex. At the bottom of the longitudinal fissure are association and commissural neurons (corpus callosum) that link the different lobes of each hemisphere and link the two hemispheres with each other.
On the surface of the cerebrum, various folds (gyri) are separated from one another by grooves (sulci), the more prominent of which are the fissures that serve as landmarks to separate each hemisphere into frontal, parietal, temporal, and occipital lobes. A central fissure separates the frontal lobe from the parietal lobe. Anterior to this fissure lay the motor and premotor cortices; posterior, the sensory and sensory cortical areas. The limbic lobe, which processes data concerning memory and emotions, is located near the medial surface of a hemisphere.
The large tracts running to and from the cerebral cortex can be classified into three major groups: the commissures, projection fibers, and association fibers.
The Commissures. The commissures are numerous nerve fibers that connect neurons of the two cerebral hemispheres, crossing from one side to the other and then to subcortical areas. Each cerebral hemisphere exerts its primary intersegmental control over the contralateral side of the body. Thus, lesions at this level are associated with a loss of function on the opposite side of the body (eg, numbness, paralysis).
The Projection Fibers. Projection fibers connect subcortical cell stations with the cortex. Many, but not all, afferent fibers reaching the cortex have their cell bodies in specific stations in the thalamus. In contrast, efferent fibers from the cortex may end in the thalamus, brain stem, or spinal cord. Projection fibers communicate with the cortex in a fan-like manner and project to the cortex in a spatial pattern so that body parts are generally represented with the foot at the superior aspect of the central fissure and the head at the temporal lobe. These primary receiving areas, however, are not the only stations of afferent pathways to the cortex.
The Association Fibers. Association fibers connect cells within the primary sensory and motor areas with other primary, secondary, and general intercommunicating sensory and motor fibers within the same hemisphere. It is by this design that protective sensory and motor responses can be quickly coordinated; eg, the head is automatically turned toward the direction of a noise or diverted from a bright light, and other appropriate somatic and endocrine actions are taken. This system, which is thought to be able to interconnect an impulse entering a single fiber indirectly with any other neuron of the body, entails 100 times as many neurons within the cortical layers as ever leave it.
The Diencephalon
The supratentorial diencephalon is that area of transition located between the cerebrum and the structures of the posterior fossa. The major structures of the diencephalon include the optic pathway, thalamus, hypothalamus, pineal body, pituitary gland, and 3rd ventricle. Each area of the cerebral cortex is closely connected to a corresponding but smaller area in the thalamus, and activation of one area causes a reciprocal activation of the other area.
Cranial Nerves I and II
The olfactory nerves (cranial I) are located at the floor of each frontal lobe, and the optic nerves (cranial II) develop as outgrowths of primitive diencephalon.
The Posterior Fossa Level
Portions of the nervous system located below the tentorium cerebelli but above the foramen magnum comprise the structures at the posterior fossa level, which are derivatives of primitive mesencephalon, metencephalon, and myelencephalon. The major structures at this level are the cerebellum, midbrain, pons, medulla, and portions of cranial nerves III XII contained within the skull.
The majority of what is called subconscious activity is coordinated and controlled in the posterior fossa area. Typical examples are blood pressure, respiration rate, body temperature, balance, and emotional patterns.
The term brain stem is a generalized term that refers to all structures of the brain except the cerebral and cerebellar hemispheres such as the medulla oblongata, midbrain, and pons. The lower 10 cranial nerves arise from the brain stem. The cerebellum, whose surface is highly convoluted with folds (folia), is a metencephalic derivation. It consists of two hemispheres and a midline vermis.
In many respects, the brain stem can be considered as the superior aspect of the spinal cord. As such, lesions in this area usually manifest as segmental and intersegmental disturbances. The segmental nerves of the brain stem are the cranial nerves, and brain stem lesions alter cranial nerve function as spinal cord lesions alter spinal nerve function. It should be noted, however, that the brain stem is an area where nerve tracts cross or have previously crossed to the contralateral side. Thus, if a posterior fossa lesion involves an intersegmental pathway, it will cause a loss of intersegmental function on the opposite side of the body. An extensive posterior fossa lesion may also affect consciousness (eg, coma).
The Longitudinal Systems
The major longitudinal systems, as classified in this manual, include the:
Cerebrospinal fluid system
Consciousness system
Motor system
Sensory system
Vascular system
Visceral system.
Lesions within these systems may occur at one or more of the horizontal levels previously described; viz, the supratentorial, posterior fossa, spinal, and peripheral levels. Symptoms of dysfunction usually arise as altered states of consciousness, incoordination, numbness, pain, paralysis, pressure, tingling, or weakness.
Basics Although Wallerian degeneration occurs within a few weeks within peripheral nerves, the process may take several months when CNS neurons are severed.
Axon regeneration is possible in a peripheral nerve if the cell body and the Schwann cells in the endoneurium are not destroyed, but such regeneration is unlikely in the CNS. In damaged peripheral nerves, the Schwann cells along the length of the degenerating fiber form myelinated tubes when they fragment, providing a cellular chain (framework) to receive axonal sprouts growing distally.
As there is no basement membrane or collagen sheaths in the CNS and oligodendroglia do not proliferate like Schwann cells, this identical process cannot occur. However, recent evidence has shown some instances of CNS axonal regeneration.
(1) muscle atrophy follows the separation of a somatic efferent, and
The function of a normal neuron is the initiation and transmission of impulses. This is an electrochemical process manifested as changes in electric potential. The metabolic activities of the neuron are only indirectly involved.
(1) electrotonic potentials (a flow of electric current) and
Electrotonic Potentials. Action potentials are initiated by an electrotonic potential, and these decremental potentials can be initiated by action, synaptic, or receptor potentials. Electrotonic potentials are the result of a voltage change, while action, synaptic, and receptor potentials are due to membrane changes.
(1) a presynaptic electric signal,
Resting Potential Cell permeability. The membrane of a cell is semipermeable; ie, it is permeable to some ions but not others. When at rest, the molecular arrangement within the neuronal membrane allows almost free flow to the small potassium ions but inhibits the diffusion of the larger sodium and calcium ions. An ion (eg, potassium) has a high conductance if it can move through a membrane easily. Thus, in terms of neuronal membrane: Potassium has a high internal ionic concentration (about 140 mEq/L), low external concentration (about 4 mEq/L), and high permeability during rest.
Sodium has a low internal ionic concentration (about 14 mEq/L), high external concentration (about 1—2 mEq/L), and low permeability during rest. Increasing the sodium concentration of the resting external medium increases the amplitude of the action spike; and, conversely, lowering the sodium concentration reduces the amplitude following stimulation.
Chloride has a low ionic internal concentration (about 8 mEq/L), high external concentration (about 108 mEq/L), and moderate passive permeability during rest.
Energy-dependent mechanisms. Certain dynamic energy-dependent metabolic mechanisms are contained within cell membrane that move ions from one side of the membrane to another so that their interior and exterior concentrations are altered.
A neuron's inactive potential is stable as long as there are adequate supplies of oxygen and glucose. However, active transport ceases and resting potential cannot be maintained if something (eg, anoxia, cold) interferes with the cell's energy-producing system or if an enzyme block occurs.
(1) ion leakage that reduces concentration gradients,
(1) A local change in applied voltage from some source (eg, electrotonic potential). It is in this manner that an action potential is initiated.
(1) the diameter of the axon (ie, the larger the axon, the lower the resistance),
Conductances carrying the inward flowing depolarizing sodium and calcium currents of the action potential are different from each other and different from those carrying the outward flowing repolarizing potassium current. Because they are separate, they can be stopped medically by certain drugs called "blockers." The strength of the stimulus. The stronger the stimulus, the more prompt the response.
The nature of the stimulus (eg, the response to sound is more prompt than that to light).
The number of synapses through which the impulses must pass; ie, a greater number delays to some extent the reaction time. The time it takes an impulse to be transmitted through a synaptic center primarily depends on the strength of the stimulus and the physiologic state of the synapse (ie, normal, hyporeactive, hyperreactive).
Synaptic Transmission
Symptoms of neurologic dysfunction may reflect a degenerative, a neoplastic, a vascular, an inflammatory, a toxic or metabolic, or a traumatic disorder.
1. By relating a patient's symptoms and signs to one of the six major longitudinal functional subsystems of the nervous system that are described in this chapter and Chapter 3; viz, the sensory, motor, visceral, vascular, consciousness, and cerebrovascular systems.
The cerebral cortex is not required for the total perception of sensation but is for its finer discrimination. Total loss of the cerebral cortex causes a "vegetative" existence. Even if large areas of the somesthetic cortex are destroyed, for example, the patient will still be able to perceive when an object touches the skin. However, the blindfolded patient will be unable to distinguish the shape, size, and other characteristics of the stimulus. Two-point discrimination will also be impaired. Likewise, destruction of the frontal lobes does not destroy a patient's ability to think. However, it does impair the patient's ability to think in abstract terms. Thus, these points (differentiation and abstraction), along with others, are commonly utilized during diagnosis to help localize the anatomical location of a lesion.
(1) sodium concentration cannot be increased or
Minor structural changes may produce an especially porous membrane that will cause increased sodium conductance, and the effect will be a partially depolarized (excitable) cell. However, if the leakage is severe, conduction will become blocked.
BIBLIOGRAPHY
The Sensory System
The afferent system receives information from the external and internal environments and transmits it to the CNS where it is processed and integrated. The major structures of the somatosensory system, from the peripheral to the central, include the receptor organs, the special sensory organs (eg, for vision, hearing, taste, smell, balance), their afferent fibers, the dorsal root ganglia, the posterior roots, the ascending tracts of the spinal cord and brain stem, portions of the thalamus, the thalamocortical radiations, and their endings in the sensory and sensory association cortex of the parietal lobe.
The Motor System
The efferent system, which is present at all major levels, governs the activity of the somatic muscles. Its major structures, from the central to the peripheral, include the motor and premotor cortices of the frontal lobe, the descending tracts that transverse the internal capsule, the cerebral peduncles and medullary pyramids, other centers in the brain stem, the descending tracts of the spinal cord, the anterior roots, the efferent fibers, and their endings within the surface membrane of skeletal muscle. The cerebellum, basal ganglia, and related pathways are also included in this system.
The Visceral System
The visceral system, which is also present at all major levels, consists of afferent and efferent neural structures that enervate the vegetative organs and homeostatic mechanisms. Its major structures, from the central to the peripheral, include supratentorial areas within the limbic lobe and hypothalamus, posterior fossa fibers of certain cranial nerves and the reticular formation, ascending and descending tracts within the brain stem and spinal cord, many ganglia and plexuses, the autonomic nerves, and their peripheral effectors and receptors.
The autonomic nervous system is responsible for the innervation of the smooth muscle of the viscera, heart, glands, and vessels. It is divided into a thoracolumbar sympathetic division and a craniosacral parasympathetic division, and these systems are generally interdependent and antagonistic in the function of their effectors.
The sympathetic system is composed of a ganglionated trunk located lateral to the vertebral column that is connected to spinal cord centers via the rami communicantes through the spinal nerves and to the viscera via the splanchnic nerves and various abdominal ganglia. The sympathetic ganglia are also connected to each other by fibers (gangliated cords). In the thoracolumbar sympathetic system, myelinated fibers communicate with the spinal cord via the white ramus. The efferent fibers leaving these ganglia (gray ramus) are essentially unmyelinated, and greatly exceed the number of preganglionic fibers. Thus, the result of excitation is quite diffuse.
The parasympathetic system consists of certain nerve fibers of some cranial and the middle sacral nerves. Cranial preganglionic fibers pass from the CNS through ventral roots but they synapse in ganglia (eg, ciliary, sphenopalatine, otic, submaxillary, sublingual) in or near the organs they enervate. Thus, the result of their excitation is more localized. Sacral fibers collect to form the pelvic nerve, which proceeds to the pelvic plexus and from which postganglionic fibers are distributed to the pelvic organs.
The Spinal Levels
Those portions of the CNS located below the foramen magnum are the spinal levels, which are surrounded by the vertebral column and sacrum. The major structures are the caudal edge of the medulla, the spinal cord, and the origins of the spinal nerves.
The spinal cord proper consists of cell bodies and fiber tracts. Cervical and lumbosacral enlargements are exhibited in the spinal cord and are related to the innervation of the upper and lower extremities. Cross sections through these enlarged areas show a relative increase in gray matter.
The spinal cord is much shorter than the vertebral canal; it ends at or near the L1 vertebral level. Thus, for most of its length, a cord segment is not on the same level as its corresponding vertebral segment. Below the L1 level, the collection of descending nerve roots within the lumbosacral canal is the cauda equina.
Bilaterally attached to the spinal cord, by anterior (motor) and posterior (sensory) nerve roots, are the 31 pairs of spinal nerves. Two exceptions to this arrangement are that C1 has only a motor root and the 1st coccyxgeal nerve (and possibly S5) has only a sensory root. As the anterior and posterior roots leave the spinal cord, they join near the intervertebral foramen (IVF) and then branch again into anterior and posterior divisions as they enter the peripheral level.
The axons of motor neurons emerge from the cord through the anterior roots to enervate voluntary muscle. They have large cell bodies that are located in the anterior gray matter of the spinal column. In contrast, cell bodies of the sensory neurons are situated in the dorsal ganglia, exterior to the CNS, and their axons enter the cord via the posterior roots and terminate in the posterior horns. Interneurons, of varying length, lie wholly within the cord. Fibers connecting the autonomic ganglia are not considered to be interneurons.
The spinal levels transmit information to and from segmental and higher central centers. Thus, a lesion at a spinal level may alter peripheral function at that level and intersegmental function below that level.
The Peripheral Levels
The peripheral levels of the nervous system include all the nervous tissue located outside the CNS; thus, the peripheral and cranial nerves and their branches. This includes the afferent and efferent somatic nerves, the autonomic ganglia and nerves, the sensory receptors, and the motor end organs.
Each pair of spinal nerve that emerges from the CNS represents a specific spinal cord segment. On their initial route to peripheral structures, nerve fibers of the larger anterior division in some areas gather in certain distributing areas (plexuses) such as the brachial plexus, which serves the upper extremities, and the lumbosacral plexus, which serves the lower extremities.
MICRONEUROPATHOLOGIC REACTIONS: GENERAL CONSIDERATIONS
Any one of the various cellular elements of the nervous system may undergo physical change as the result of trauma or dysfunction. Such structural changes may reflect damage produced by a pathologic process or the reaction to it. Pathologic changes may also be primarily functional with little change in structure. Such changes at the spinal and peripheral levels are a primary concern within chiropractic and serve as the basis for many adjustive and reflex therapeutic approaches.
In many pathologic conditions, the various cells react in concert and their reflected clinical features are nonspecific. For example, CNS neuronal loss is invariably accompanied by loss of astrocytes and microglia at the site. At other times, specific changes occur that show or suggest a particular disease entity.
Cerebral Edema
Cerebral edema may be caused by a large variety of factors. It is the direct effect of fluid accumulating in the brain, especially in its white matter, and may be diffuse around or localized within a focus of injured tissue. Astrocytes, myelin lamellae, perivascular spaces, and extracellular spaces may be swollen, depending upon the pathophysiologic mechanisms involved.
Neuronal Pathology
Ischemic Cell Changes. Oxygen may be deprived from a nerve cell by anoxia, lack of circulating blood, vasospasm, embolism, lack of metabolic substrates, or the presence of poisons that block oxidative metabolism.
When oxygen is deprived from a nerve cell, oxidative metabolism ceases. The immediate physiologic change is an acute swelling of the neuron. Subsequently, in a matter of hours (8—12), irreversible changes occur: the neuron becomes shrunken, the cytoplasm becomes diffusely eosinophilic, the nucleus becomes pyknotic, and the outline of the cell body becomes sharply angular. Complete dissolution of the cell follows these morphologic changes.
Central Chromatolysis
This is an axonal reaction that often occurs especially in large motor cells within the anterior horns of the cord or motor nuclei of the brain when their axons are damaged proximally. The cell bodies swell, their Nissl bodies dissolve, and the nucleus migrates peripherally. This process starts in 2 or 3 days after injury and reaches maximum in 2—4 weeks. However, unlike the results of ischemia, central chromatolysis is reversible within a few months if the cause has been removed.
Wallerian Degeneration
In contrast to ischemic effects and central chromatolysis, Wallerian degeneration begins at the distal aspect of the axon after its cell body has been separated from the axon by trauma or disease. As an axon is nourished primarily by axoplasmic flow from the cell body, a severed axon cannot survive. The neurofibrils disappear rapidly, then the cylinder of the fiber breaks up into short fragments, and finally the fragments are dissolved (apparently digested by chemicals from its myelin envelope).
The process of Wallerian degenerative is not identical in the peripheral and central nervous systems:
Wallerian degeneration does not usually extend postsynaptically. There are, however, two important exceptions to this rule:
(2) certain CNS pathways can be affected by transneuronal degeneration. Why this occurs has not been determined, but it is thought that presynaptic fibers exert some type of postsynaptic trophic influence.
Storage (Balloon) Cells
Many metabolic diseases affecting the nervous system are associated with an accumulation of substances. Tay-Sachs disease (lipid storage disease) is an example. As the substances accumulate, the cytoplasm of the involved cell bodies become severely swollen, and the nucleus and Nissl bodies are forced to a corner of the angular cell. The cause can only be identified by biochemical (usually postmortem) identification of the accumulating material.
Inclusion Bodies
Inclusion bodies are discrete deposits within nerve cells whose presence often identifies a specific type of disease process. For example, they appear in the cytoplasm as Lafora bodies in myoclonus epilepsy, Lewy bodies in Parkinson's disease, Negri bodies in rabies, and Pick bodies in Pick's disease. They also appear in the nucleus as Cowdry type A inclusions in various viral infections.
Neurofibrillary Degeneration
Neurofibrillary degeneration is a process in which masses of neurofibrils become clumped within the cytoplasm of CNS cells. It is a type of neuronal degeneration that is closely related to advanced age but is especially prominent in clinical dementia (Alzheimer's neurofibrillary degeneration). Senile plaques are usually associated, which consist of a central core of amyloid deposits surrounded by a ring of degenerated axons, astrocytes, and microglia.
Oligodendroglia and Schwann Cell Pathology
Oligodendroglia and the myelin sheaths they create in the CNS are highly sensitive to injury, and when damaged, their nuclei rapidly fragment and dissolve. In demyelinating (myelinoclastic) diseases (eg, multiple sclerosis), the myelin appears to be attacked by some unknown substance that breaks the myelin into its component lipids, after which they are absorbed. Rarer myelin diseases are the genetic dysmyelinating (leukodystrophy) disorders, where the myelin is defectively formed and unstable due to a metabolic error.
Disease processes affecting Schwann cells are associated with segmental myelin breakdown and loss (demyelination). If the process is severe, the axon becomes affected and Wallerian degeneration occurs. In certain rare genetic disorders of peripheral nerves (hypertrophic neuropathy), nerve fibers undergo repetitious demyelination and remyelination. Each episode leaves a thin film of scar tissue in concentric layers, the axons thus progressively become larger and firmer until they are destroyed, and then only the connective tissue remains.
Microglia Pathology
Microglia react predictably in most diseases of the CNS. Microglia cells near the site of irritation greatly multiply, and their nuclei elongate to form rod cells. The process may be halted here in low-grade disorders such as chronic syphilitic encephalitis (general paresis). If necrosis continues, the cells become phagocytic and engulf necrotic debris and damaged neuron fragments in the area involved.
Astrocyte Proliferation
Astrocytosis (gliosis), the effect of almost any injury of the CNS, is the formation of scar-like changes within injured neural tissue. During the reaction, astrocytes greatly increase in number and progressively form longer and thicker processes that form a dense network within and around the injured area.
In certain CNS diseases, especially metabolic disturbances, the nuclei of astrocytes undergo morphologic changes. Alzheimer's disease and hepatic failure are typical examples. In certain CNS viral diseases, intranuclear inclusion bodies may develop within astrocytes.
When noxious stimuli are present (eg, infection) in the ventricular system, ependymal cells are readily destroyed, subependymal astrocytes proliferate, ependymal granulations develop, and the tissue becomes infiltrated with leukocytes.
NEURONAL ELECTRONEUROPHYSIOLOGY
Basic Considerations
All nerve and muscle cells have various electric potentials. In nerve fibers, these include resting (inactive) potentials, active potentials, and various local potentials (viz, generator, electrotonic, synaptic).
Membrane Potentials
In living tissues, current is caused by the movement of charged ions, and it can flow through the low-resistant neuronal membrane as the result of an applied voltage or a change in membrane conductance. The difference in the electric charge between the inside and outside of the cell is the membrane potential. When a nerve cell is inactive, its baseline level is its resting potential and determined primarily by potassium ion concentrations.
The cytoplasm of a neuron is a highly conductive electrolytic solution, and any change in the potential of one part produces an equal change in the potential at all other points. Thus, any local change in membrane potential will produce a flow of current throughout the cell by initiating an action potential. When impulse activity exists, action potentials are produced and operate according to the all-or-none principle in the neuron, spread to all parts of the cell, and are determined primarily by sodium ions.
Local Potentials
Localized changes in membrane potential that are the reaction to a stimulus are called local potentials. All local potentials are graded potentials (ie, their amplitudes are proportional to the size of the stimulus); thus, their strength is proportional to the strength of the stimulus. Unlike action potentials, local potentials remain localized in the area of the cell irritated, can be summated and integrated within single neurons, and do not operate according to the all-or-none principle.
Local potentials are generated by
(2) changes in the characteristics of resting receptor membranes or synapses.
Receptor (Generator) Potentials. Receptors are specialized to respond to certain types of stimuli (eg, light, sound, touch, heat, etc). Once stimulated, they generate local electrotonic potentials that, in turn, initiate action potentials. It should be noted that sufficient depolarization from any origin anywhere on the axon can initiate an action potential.
In sensory neurons, however, it is usually started at a membrane (receptor) distal to the cell body; in motor neurons, it is usually started by synaptic potentials near the axon hillock (generator). It takes about 30 millivolts of excitatory postsynaptic potential to elicit an action potential in a nerve cell body but only 11 millivolts to achieve an action potential at the hillock area.
Synaptic Potentials. These are changes in potentials that occur in postsynaptic cell membranes in response to chemical or electronic message transmission produced presynaptically. They usually involve an electrochemical and chemoelectric conversion process; ie,
(2) a synaptic chemical signal, and
(3) a postsynaptic electric signal.
Extracellular fluid contains positively charged ions (anions) of sodium, potassium, and calcium, and negatively charged ions of chloride (cations). Intracellular fluid contains many negatively charged indiffusible ions, which are derived from protein. If a voltage is applied, these anions and cations move towards a pole with an opposite charge.
A nerve or muscle cell's resting potential is in the range of 70 to 90 millivolts. The sensitivity to neuronal membrane becomes greater as the voltage becomes more negative and less sensitive as the voltage rises. Resting potential is determined by two factors:
The steady resting potential of a neuron is presently thought to be a balance struck among
(2) ion pumping that increases concentration gradients, and
(3) the cell's internal negative charge that repels anions and attracts cations.
The Sodium Pump
Nerve cells obtain energy from oxygen and glucose through glycolytic pathways, the Krebs cycle, and the electron transport system all of which provide the energy necessary for neuronal function in the form of adenosine triphosphate (ATP). ATP, provided by the mitochondria, is important during resting potential for the system (active transport) that moves potassium into and sodium out of the nerve cell. This hypothetical mechanism within the neuronal membrane (sodium pump) allows this movement against their concentration gradients, and it is not confined to nerve cells.
The movement of ions across a permeable membrane occurs freely by diffusion if the electrochemical gradient does not oppose it. In the recovery phase following an action potential, the outward flow of sodium has to be forced against a gradient. This is the basis for the "sodium pump" theory, and the energy to maintain this pump is thought to be derived from metabolic processes within the nerve. Prichard states that it is thought to consume 20% of the entire energy requirements of the animal body.
It is known that the result of active transport through the cell membrane is an unequal concentration of ions on opposite sides of the membrane. Consequently, there is a small continuous efflux of potassium ions and an influx of sodium ions, but these ions are transferred back through the membrane by the sodium pump to maintain a dynamic steady resting state rather than a true state of equilibrium.
As a large number of positive sodium ions are pumped to the exterior, this leaves a large number of nondiffusible negatively charged ions internally (eg, protein ions, phosphate ions, etc). Thus, the interior surface of the resting membrane becomes negative and the exterior surface becomes positive in charge.
The sodium pump appears to be a membrane enzyme that is capable of splitting ATP and using the energy released to move potassium and sodium ions through the membrane against their concentration gradients. This system is also thought to be a "coupling system" in that, if either ion is removed from the system, the remaining ion is not transported well.
A hypothetical sodium/calcium countertransport pump, a chloride pump, and an ATP-requiring calcium pump have been brought forth in addition to the ATP-requiring sodium/potassium pump. Absolute proof of the existence of such pumps has not been established, but the theories explain most experimental findings.
Local Effects
Local effects are the result of a shift of membrane potential within a specific area of the cell due to a hyperpolarizing or depolarizing stimulus. This may be an effect of:
(2) A local change in membrane permeability to one or more ions. This occurs in response to neurotransmitters (synaptic potential) or external stimuli (receptor potential) and is the mechanism by which signals are transmitted to or from the neuron.
Neuronal Summation
It was previously mentioned that all local potentials are graded potentials. These changes in amplitude rise and fall occur over several milliseconds, far outlasting the stimulation time. If a second local stimulus is applied to a neuron before the effect of one has subsided, the result is a larger local potential. Thus, local potentials can be summated. It is in this manner that individual stimuli that are below threshold can be increased in value collectively to produce an action potential.
Temporal summation is achieved because local potential changes initiated close to each other in time are additive. Spatial summation is achieved in border zones when local potential changes are initiated at different sites. Thus, through temporal and spatial summation, the cell body can integrate stimuli from different sources that converge on an area of cell membrane.
Neuronal Accommodation
Changes occur in the ionic conductivity of neuronal membrane whenever a local stimulus is applied for more than a few milliseconds. These changes tend to restore the membrane to its inactive baseline potential even if the stimulation is continued. This phenomenon (accommodation) can occur if a stimulus is increased slowly, and the effect is no change in membrane potential.
Action Potentials
A neuron's inactive potential can be changed by either the membrane becoming less negative outside (depolarization) or more negative inside (hyperpolarization). Rapid transient changes occur in response to electrical, mechanical, or chemical stimuli; while prolonged changes, which may last for many minutes, result from anoxia or effects that change the concentration of ions on either side of the membrane.
Excitability
A neuron's minimal level of excitability is determined by its threshold, which may be reached either by a single sufficient stimulus or by summation. The term threshold is used in two senses. First, it refers to the voltage required to induce an action potential such as when extrinsic stimulation is applied and recorded. Second, it is used to describe the relative state of membrane potential. For example, the closer the membrane potential is to threshold, the easier it is to fire; the farther the membrane potential is from threshold, the more difficult it is to fire.
The first portion of the membrane potential change is that of the local potential. The rapidly rising phase of the action potential occurs at threshold as an effect of the influx of positive ions (sodium and some calcium). During long intervals between action potentials, the sodium pump continually transports internal sodium to the exterior.
Under typical conditions, the threshold of a neuron for excitation is 59 millivolts, representing a postsynaptic excitatory potential of +11 millivolts; ie, 11 millivolts more positive than a neuron's average resting potential of 70 millivolts. Once the threshold is reached, the membrane becomes suddenly more permeable to sodium, which is thought to be related to a change in the binding of calcium in the membrane. This shifts the membrane potential toward the equilibrium potential of sodium rather than that of potassium. The polarity of the membrane is reversed by this depolarization, and a flow of current occurs as many sodium ions and some calcium ions flow inwardly and the abundant potassium ions, subsequently, flow outwardly. This change in conductance results in the action potential.
At the phase when sodium conductance is high internally, a stimulus applied at this time will have no effect; ie, action potentials, unlike local potentials, cannot be summated. This period of complete unresponsiveness (absolute refractory period) is followed by the relative refractory period, as sodium conductance begins to return to its resting equilibrium, where a greater stimulus will produce a smaller action potential.
Propagation
Besides the horizontal current produced by the flow of positive sodium ions through an area of depolarized membrane, there is also a longitudinal current (electrotonus), which, unlike the action potential, has no refractory period. It flows on the interior and exterior surfaces of the membrane and tends to depolarize the areas surrounding the action potential.
This depolarization (an electrotonic potential) is usually sufficient to shift the membrane potential to threshold and produce an action potential in the adjacent area of membrane. In this manner, the action potential progresses away from the site of initial stimulation along the axon. The potential is prevented from reversing backward into an area just depolarized because of the refractory period. That is, after passage of a nerve impulse, repolarization and ionic balance during the refractory period are quickly restored so that another impulse can be received.
There are several other theories regarding the electric propagation of the nerve impulse. However, all recognize a decrease in resistance across the membrane as a crucial event for the impulse to pass. The degree of resistance may be regarded as an index of the membrane's permeability to certain ions. However, unlike a membrane's resistance, the capacitance of nerve membrane does not change appreciably during the passage of an impulse.
The conduction rate of the action potential is determined by
(2) the quantity of current necessary to produce depolarization in adjacent membrane, and
(3) whether the axon is myelinated. In myelinated nerves, the active region "triggers" the resting area ahead only at the nodes of Ranvier, where myelin is absent; ie, the nerve impulse is propagated from node to node in a leaping manner (salatory transmission). This occurs because an action potential at one node produces sufficient longitudinal current to depolarize the next node.
Facilitation
After repetition of a reflex act, its establishment becomes increasingly easy, and, within limits, the reaction time lessens. This type of facilitation is the basis of habit formation. The term facilitation is also used to describe the state of a neuron that has been summated to some degree below its firing threshold at which point meager stimuli from some source can easily excite the neuron.
Reaction Time
The time elapsing between the application of a stimulus and the beginning of a response is the reaction time. Reaction time varies in different individuals and in the same individual under different conditions. The most commonly involved factors are:
Membrane depolarization caused by an action potential on presynaptic terminals causes the vesicles to empty, and the quantity of neurotransmitter released at a presynaptic vesicle is directly proportional to the change in membrane potential (ie, its amplitude).
Calcium flows into the terminals when the depolarization alters its conductance, and this appears to trigger the release of the neurotransmitter, which diffuses across the synaptic cleft. Sodium and magnesium ions are also involved in this triggering process. The neurotransmitter extruded into the cleft excites or inhibits the subsequent neuron depending upon the type of receptor contacted.
Once an excitatory neurotransmitter contacts the specialized cells of the postsynaptic membrane, the membrane's permeability is altered, the local potential is changed, ionic conductance is altered, sodium ions enter the cell, and a flow of current is produced. This phase of sodium permeability lasts for 1—2 milliseconds, but the potential may persist for about 15 milliseconds until the sodium pump becomes effective in re-establishing the resting potential.
The synaptic potential, like a receptor potential, is a graded response that initiates a local electrotonic potential. The neurotransmitter may also act on a second chemical messenger in the postsynaptic membrane to create a longer change in permeability via an effect on the sodium pump.
Synaptic Inhibition
With a few exceptions, each neuron releases the same type of transmitter substance at all its terminals (Dale-Feldberg law) and that substance will have either an excitatory or inhibitory influence on all associated postsynaptic receptors relative to their properties and local hormonal influences. As postsynaptic effects are receptor dependent, a neuron releasing the same transmitter may cause an inhibitory effect on some connecting neurons and an excitatory effect on other connecting neurons.
Some axon end feet within the CNS release inhibitory transmitters. Common examples are the amino acid glycine and GABA (gamma aminobutyric acid). Inhibitory neurons tend to block unimportant signals and thus allow transmission of selected messages (eg, to centers that otherwise would be subjected to a continuous bombardment of impulses from receptors located throughout the body). Inhibition can also occur if mechanisms produce a postsynaptic hyperpolarization that moves the membrane potential farther from its threshold.
Presynaptic Inhibition
Because the release of a neurotransmitter is a graded response governed by the amplitude of the action potential at the vesicle, changes in membrane potential at the terminal will modify synaptic transmission. This is typically seen where some terminals synapse directly on others that they can partially depolarize and thus inhibit release when they are acted upon by an action potential.
The most common occurrence is seen in peripheral synapses of sensory paths. The result (presynaptic inhibition), therefore, is a smaller release, and this effect is usually of a longer duration (eg, 0.5 sec) than is postsynaptic inhibition (eg, 10—15 milliseconds). The same timeframe is true for excitation.
Postsynaptic Summation
When either peripheral or central impulses from several sources act on single cells, synaptic potentials will summate both spatially and temporally.
A single end foot has been computed to cause an excitatory postsynaptic potential of about 1 millivolt or less. As the usual threshold for excitation is 10 or more millivolts, a minimum of ten end feet firing simultaneously anywhere on a receptive membrane of a nerve cell, even at widely separated areas, is necessary to produce an action potential. This is an example of postsynaptic spatial summation, and it occurs because a potential change at any single point within the cytoplasm will cause the potential to change throughout the cytoplasm.
In addition, as each end foot can fire at intervals about 1 millisecond apart and its postsynaptic effect may last up to 15 milliseconds, a rapidly firing end foot will have a summed postsynaptic effect. This is an example of postsynaptic temporal summation. Through either spatial or temporal summation, it can be seen that potentials derived from presynaptic, synaptic, or postsynaptic excitatory and inhibitory mechanisms of identical value will cancel each other.
CLINICAL APPLICATIONS
Degenerative disorders are characterized by gradual but widespread neuronal damage that manifest as diffuse, chronic, progressive diseases. As any cell within the nervous system can undergo neoplastic changes, neoplastic disorders feature one or more gradually enlarging masses of proliferating cells. The type of neoplasm is named after the predominant cell type (eg, astrocytoma, oligodendroglioma, Schwannoma).
Vascular disorders are invariably associated with acute symptoms and signs that may be focal or diffuse, depending upon cause and site (eg, hemorrhage, anoxia, encephalopathy). Inflammatory disorders usually exhibit progressive subacute symptoms that may be focal (eg, abscess) or diffuse (eg, encephalitis, meningitis). Toxic or metabolic disorders are characterized by widespread neural dysfunction exhibiting diffuse signs and symptoms that may arise as an acute, a subacute, or a chronic process.
Traumatic disorders affecting the nervous system as usually focal and have an acute onset, reflecting the offending agent and the area of tissue damaged. However, delayed effects may produce signs and symptoms pointing to a pattern of progressive degeneration.
Importance of Normal Findings
In nerve, muscle, or gross static and dynamic analyses or any other diagnostic procedure, a clinician may use a normal finding to eliminate certain disorders during the differential diagnosis process. When properly interpreted, however, a normal value may also aid in differentiating among possible diagnoses that yield normal values with different frequencies in occurrence.
Gorry and associates suggest a simple method that has the examiner estimate the probability of various diagnoses and then combine these estimates with the anticipated frequency of negative results for each disorder under consideration. Thus, from a clinical standpoint apart from the medicolegal factor involved, the recording and appraisal of a normal finding can be just as important as that of an abnormal finding.
Applied Neuroembryology
Developing cells change their response to metabolic injury as they grow, and the embryo changes metabolically as it develops. Although developmental patterns are determined primarily by genetics and latent genetic defects may be precipitated by noxious agents (eg, anoxia, hypoglycemia, drugs, radiation, toxins), different agents may produce different types of malformations at the same stage of development.
Most gross embryologic malformations such as meningocele and meningomyelocele are rarely seen in the chiropractic office. However, various anomalies not obvious externally are often shown in roentgenography for another reason (eg, spina bifida occulta, hemivertebrae, etc). Apart from such developmental defects, knowledge of neuroembryology is helpful in localizing the site of neural damage and in recognizing various patterns of pain.
Daube/Sandok explain that because the nervous system and the skin both derive from ectoderm, a genetically determined disorder of one system may affect the other. This is often seen in the neurocutaneous syndromes and ectodermal dysplasias. For example, in Sturge-Weber syndrome, meningeal angioma and cortical calcification manifest in the nervous system and a port-wine nevus manifests on the face. In tubercular sclerosis, cortical giant cell tumors, gliomas, and periventricular tumors arise in the nervous system and depigmented patches, subungual fibromas, and adenoma sebaceum appear in the skin. In von Recklinghausen's disease, schwannomas and gliomas appear in the nervous system and neurofibromas and caf-au-lait spots arise in the skin.
The CNS and the retina of the eye are also closely related embryologically, and developmental syndromes often manifest both CNS and retinal signs. Typical examples are the retinal tumors associated with neuroectodermal dysplasia and the cherry-red macula spot of Tay-Sachs disease.
Pathoanatomical Correlations
Rarely do patients enter a doctor's office with a singular complaint. They invariably mention a few symptoms that are a priority concern to them, and, upon questioning, will reveal several associated concerns. The clinician will then use inductive and deductive reasoning to arrive at an initial picture that will be constantly altered, refined, and reorganized according to the findings of subsequent examinations and patient progress. With knowledge of the neuroanatomy and neurophysiology of the various parts of the nervous system, neurologic signs and symptoms can be related to specific areas of distress within the nervous system in most instances.
Topographic Localization
The term localization means determining the location along various pathways where function is interrupted. It becomes precise when one or more areas where both segmental and intersegmental functions are altered in one or more systems and they can be identified.
During the diagnostic process, the topographic localization of a neural deficit is arrived at by two basic steps (dimensional viewpoints):
2. By localizing the lesion at one of the four well-defined horizontal levels of the nervous system described in this chapter and Chapter 4; viz, the supratentorial, posterior fossa, spinal, and peripheral levels. These levels are the points at which the function of each longitudinal system can be integrated and modified.
Diffuse vs Focal Lesions
If the responsible lesion is diffuse (involving bilateral, usually symmetrical, areas but not extending across the midline), its anatomical level(s) should be noted. A diffuse lesion may involve a single level or be distributed over multiple levels. If the lesion is focal (well-circumscribed), its anatomical level should be noted as well as whether it is located on the left, right, or midline of the body. Note that a focal lesion may involve the midline and contiguous structures bilaterally. The term multifocal refers to lesions involving more than one circumscribed area or several noncontiguous structures.
The history of a neurologic disorder's onset and progress is usually extremely helpful in its diagnosis. Diffuse disorders may be acute, subacute, or chronic. Diffuse vascular disorders (eg, hemorrhage, anoxia) are usually acute; inflammatory disorders (eg, meningitis, encephalitis), subacute; and degenerative disorders, chronic. Toxic and metabolic disorders are usually diffuse; their timeframes will depend upon the specific toxin or metabolite involved.
Almost all progressive focal lesions are space-occupying lesions (eg, hemorrhages, abscesses, tumors), and the specific type of mass determines its course. Focal vascular disorders (eg, hematoma, infarct) are usually acute; inflammatory disorders (eg, abscess), subacute; and neoplasms, chronic. Traumatic disorders usually manifest in an acute manner, but their specific localization and course are determined by their site, severity, and the complications involved.
Transient Physiologic Disorders
Neurologic disorders with a history of transient episodes, lasting from seconds to hours, are often seen in chiropractic practice. Their cause can generally be attributed to temporary neuronal hyper- or hypo-activity, and a few typical examples will be described in this section. Symptom manifestation will vary depending upon which specific sensory or motor cells are involved; eg, twitchings, hyperesthesia, photophobia, abnormal behavior, muscle weakness, numbness, or tingling.
Potassium Imbalance
A neuron's resting potential is primarily determined on the internal and external concentrations of potassium. For example, an increase in extracellular potassium will produce a resting potential closer to threshold (ie, a partially depolarized neuron); thus, the cell will become so sensitive that it will fire upon normally subthreshold stimuli. In near-threshold levels of extracellular potassium, the cell may fire spontaneously. This may be the direct cause of idiopathic pain or the convulsions associated with transient hypoxia. In very high levels of extracellular potassium with an extremely low concentration gradient, however, firing function is invariably lost.
A reduction of extracellular potassium will hyperpolarize the resting potential, and the neuron will become less excitable because the membrane potential is farther from threshold value. This effect is commonly seen when strong diuretics are prescribed (eg, for hypertension, obesity, PMS) and is typical of hypokalemic periodic paralysis and some renal diseases.
Sodium Imbalance
A neuron's action potential is primarily determined on the internal and external concentrations of sodium. Imbalances can be an effect of alterations in sodium conductance or ion concentration. For example, a membrane cannot conduct an action potential if
(2) conductance is increased far beyond normal.
Calcium Imbalance
A reduction in extracellular calcium causes decreased concentration gradients for both potassium and sodium. Thus, calcium serves as a membrane stabilizer. In hypocalcemia, for example, it is common to see increased neuronal excitability and sometimes spontaneous activity because of the reduced resting potential. This state will usually manifest as paresthesias or involuntary muscular twitching. However, an opposite effect may be seen because an extremely low calcium level will block synaptic transmission.
An increase in extracellular calcium, rarely demonstrable overtly when mild or moderate, has the opposite effect. That is, action potential will be blocked, but synaptic transmission will be enhanced.
Drug Reactions
Certain drugs may alter normal function of the axon or synapse (presynaptically or postsynaptically). Local anesthetics usually prevent changes in sodium conductance within the axon's membrane to the degree that action potentials become blocked. At the synapse, transmitter release may be inhibited or postsynaptic receptors may be blocked by certain drugs.
Nerve Fatigue
The effect of chronic pain on the nervous system brings about changes in the nervous system similar to those produced by prolonged muscular effort. Prolonged pain, however, is not the only mechanism that can cause such an effect. The onset of clinical fatigue appears to be favored by poor health, postural stress, chronic aches and discomforts, mental/emotional conflicts, and periods of intensively prolonged concentration or eyestrain.
Work done under compulsion (eg, a sense of duty) results in nervous system fatigue more readily than when interest and motivation are high. Worry and fear have long been known to cause effects within the autonomic nervous system. It is for this reason that indigestion, palpitation, hyperhidrosis, constipation, and vasomotor headaches are commonly associated with neurasthenic conditions.
Similar effects as those seen in neuronal fatigue occur at synapses as the result of the accumulation of circulating metabolic waste products or fatigue-producing substances developed within the brain and in toxicosis. They are also seen in some psychic disturbances such as depression and after prolonged concentration.
The higher cerebral centers are most easily and quickly affected. It is thought by many authorities that it is for this reason that young children "go stale" quickly unless their mental training is properly balanced by rest and recreation. The same effect is likely seen in adult "burnout."
It is often stated that change of activity is equivalent to rest. It is questioned whether this is true of mental activity, but it may be true of muscular activity. The explanation offered by Kimber and associates is that when nervous stimuli are altered, new pathways and groups of muscles are called into play. For example, a person who is tired of inside work may go outside and walk, because the fresh air, changed environment, and different thoughts serve as new stimuli, travel different CNS paths, and throw into activity different groups of muscles.
The fatigue of nerve cells resulting from typical daily activities is normally repaired by a night's sleep. However, if unnecessarily demanding activities are continued day after day for an extended period, the effects of fatigue accumulate and a pathologic state is likely to result. This is more apt to happen if mental conflicts add their quota.
If monotony is also added (a situation where the same neurons are activated by similar stimuli), abnormal states variously known as brain fag, burnout, neurasthenia, or a nervous breakdown are likely to develop. Depression or exhaustion of the higher CNS centers is usually the starting point of such a condition.
If motivation or will-power is weakened, the power of concentration lessened, and the mental faculties impaired, the imagination is likely to substitute morbid ideas, fears, and phobias that under ordinary circumstances would not arise, or, if they did, they would be quickly dismissed or reasoned away.
Neural Regeneration
After pressure injuries or toxic neuropathy, peripheral nerves have great recuperative potential. Unfortunately, expectations are usually disappointing following trauma or severe pathology.
Because the cell body is essential for the nutrition of the whole cell, it follows that if the processes of a neuron are severed by trauma or disease, they will die from malnutrition. If, for instance, a postganglionic spinal nerve is cut, all portions distal to the cut will die because the processes have separated from their cell bodies in the ganglion or spinal cord. The affected axis cylinder fragments, the axon breaks up, and Schwann cells proliferate to form digestive chambers. The fibers and the medullary sheath degenerate and are absorbed, and, in time, an endoneurial tubule (band of Buengner) exists that is completely filled with Schwann cells. Endoneurial tubes do not degenerate as axons and myelin do after injury. Because they are filled with proliferated Schwann cells, they remain available for entry by regenerating axons.
Segments of the proximal axons develop sprouts that tend to grow along the course of the degenerated distal segments. This appears to be a random process that is guided somewhat by the endoneurial tubules. Some succeed in uniting with healthy segments if they enter a tubule and others do not. If nerve compression is relieved, additional sprouts are encouraged to grow.
A destroyed CNS nerve, in contrast, does not necessarily mean that function cannot be restored to a considerable extent. Illis explains that previously unused or little used pathways may become effective through neuromuscular education following degeneration of major connections.
Pressure Neuropathy
Prichard points out that nerves are richly supplied with anastamoses in the longitudinal direction. This allows nerves to be entirely freed from surrounding tissues for lengths up to several centimeters without altering their function. "What nerves cannot stand is pressure and stretching; these lead rapidly to functional deterioration, which eventually becomes irreversible and ends in physical degeneration if the pathologic condition is not relieved." For this reason a surgeon cannot stretch a cut nerve to fill a widely separated gap. Approximation must be achieved by relocating the nerve's course, thus sacrificing some small branches, or by a nerve graft.
Pressure on a nerve interferes with the circulation to the neuronal membrane and with its axoplasmic flow. It is theoretically possible that pressure, if severe enough, can directly block conduction, but this would be a rare circumstance. A local block can occur within a few minutes after pressure is applied, but this is the effect of total ischemia. Even in cases of chronic partial ischemia, conduction is reduced as it passes through the affected area. In such situations, the adverse effects of any degree of ischemia may be cumulative and eventually lead to a complete conduction block.
Interference with axoplasmic flow by direct pressure or the associated ischemia will also produce functional failure in a matter of hours. If this pressure is removed, recovery will occur quite rapidly depending upon how fast the effects of the metabolic damage can be repaired. If the pressure is not removed, more serious peripheral degeneration effects occur in the neuron.
Traumatic Neuropathy
The connective tissue ends of a sharply severed nerve readily unite by cicatrical tissue while the fibers fragment and are absorbed. Before healthy fibers can unite during the regeneration process, they must, in some manner, pass over or around the intervening scar tissue. If they cannot, they may form a tangled mass of neural and connective-tissue elements (neuroma).
In most traumatic situations, there is often considerable local edema and tissue reactions around the injured neurons to offer pressure complications besides the typical processes of Wallerian degeneration.
Toxic Neuropathy
The last stages of toxic neuropathy resemble the typical Wallerian degeneration that follows nerve trauma except that the changes are scattered along the length of the nerve rather than being concentrated at a focal area. Various complexities arise if the disease process is specific for myelin, axons, or nerve cells.
Adams RD, Victor M:
Principles of Neurology, ed 2.
New York, McGraw-Hill, 1981.
Adrian ED:
Sensory mechanisms: introduction.
In Field J, Magoun HW (eds): Handbook of Physiology.
Washington, DC, American Physiological Society, vol 1, 1959.
Adrian ED:
The all-or-none principle in nerve.
Journal of Physiology, 47:460— 474, 1914.
Afifi AK, Bergmann RA:
Basic Neuroscience.
Baltimore, Urban & Schwarzenberg, 1980.
American Chiropractic Association:
Chiropractic State of the Art. Arlington, VA, 1984, pp 8—9.
Baker AB (ed):
Clinical Neurology, ed 2. New York,
Harper & Brothers, Hoeber Medical Division, 1962.
Basmajian JV (ed):
Grant's Method of Anatomy, ed 9.
Baltimore, Williams & Wilkins, 1975, pp 39-49, 479-492.
Bennett MVL:
Electrical transmission: A functional analysis and comparison with chemical transmission.
In Brookhart JM, Mountcastle VB (eds): Handbook of Physiology.
Baltimore, Williams & Wilkins, pp 357-358.
Biederman F:
The Fundamentals of Chiropractic from the Perspective of a Medical Doctor.
Heidelberg, Haug, 1968.
Bishop B:
Neural plasticity.
Physical Therapy, part 1, 62(8):132-134; part 2, 62(9):1275-1282, 1982,
Bittner H, Harper WD, Homewood AE, Janse J, Weiant CW:
Chiropractic of today.
ACA Journal of Chiropractic, 7(11):S81-88, 1973.
Brazier MAB:
Electrical Activity of the Nervous System, ed 4.
Baltimore, Williams & Wilkins, 1977, pp 1-91.
Brunarski DJ:
Clinical trials of spinal manipulation: A critical appraisal and review of the literature.
Journal of Manipulative and Physiological Therapeutics, 7(4):343-347, December 1984.
Bullock TH:
The neuron doctrine and electrophysiology.
Science, 129:17, 1959.
Burke DC, Murray DD:
Handbook of Spinal Cord Medicine.
London, Macmillan, 1975, pp 1-12.
Chusid JG:
Correlative Neuroanatomy & Functional Neurology, ed 19.
Los Altos, CA, Lange Medical, 1985, pp 82-86, 322-337, 473-492.
Clark RG:
Manter and Gatz's Essentials of Clinical Neuroanatomy and Neurophysiology, ed 5.
Philadelphia, F.A. Davis, 1975.
Coers C, Woolf AL:
Innervation of Muscle.
Springfield, IL, Charles C. Thomas, 1959.
Curtis DR, Eccles JC:
Synaptic action during and after repetitive stimulation.
Journal of Physiology (British), 150:374, 1960.
Daube JR, Sandok BA:
Medical Neurosciences.
Boston, Little, Brown, 1978, pp 2-4, 8-21, 24-44, 46-67, 68-89.
Eccles JC:
The synapse.
Scientific American, 212(1):56, 1965.
Everett NB:
Functional Neuroanatomy, ed 6.
Philadelphia, Lea & Febiger, 1971.
Faulk WP, Demaeyer EM, Davies AJ:
Some effects of malnutrition on the immune response in man.
American Journal of Clinical Nutrition, 27:638-646, 1974.
Gardner EB:
The neurophysiological basis of motor learning: a review.
Journal of the American Physical Therapy Association, 47:1115, 1967.
Glynn IM, Karlish SJD:
The sodium pump.
Annual Review of Physiology, 37:13-55, 1975.
Gordienko AN (ed):
Control of Immunogenesis by the Nervous System.
Washington, DC, US Department of Commerce, 1958, pp 22-33, 45-52, 53-58.
Gorry GA, Pauker SG, Schwartz WB:
The diagnostic importance of the normal finding.
New England Journal of Medicine, 298:486-489, 1978.
Granit R:
The Basis of Motor Control.
New York, Academic Press, 1970.
Guyton AC:
Textbook of Medical Physiology, ed 6.
Philadelphia, W.B. Saunders, 1981, pp 560-576.
Guze H:
The role of chiropractic in an integrative psychotherapy.
In Schwartz HS (ed): Mental Health and Chiropractic, A Multidisciplinary Approach.
New Hyde Park, NY, Sessions, 1973, pp 148-154.
Haldeman S, Hammerich K:
The evolution of neurology and the concept of chiropractic.
ACA Journal of Chiropractic, 7(8):S57-63, 1973.
Haldeman S:
Interactions between the somatic and visceral nervous systems.
ACA Journal of Chiropractic, V:57-64, August 1971.
Haldeman S:
Neurophysiological concept of importance to chiropractors.
Journal of the Canadian Chiropractic Association, 18(3):6, 1974.
Harper WD:
Anything Can Cause Anything, ed 3. Seabrook, TX,
published by author, 1974.
Haymaker W (ed):
Bing's Local Diagnosis in Neurological Diseases, ed 14.
St. Louis, C.V. Mosby, 1956.
Hensinger RN, MacEwen GD:
Congenital anomalies of the spine.
In Rothman RH, Simeone FA (eds): The Spine.
Philadelphia, W.B. Saunders, 1975, vol I.
Hoag JM:
The musculoskeletal system: A major factor in maintaining homeostasis.
Journal of the American Osteopathic Association, 78:562, 1979.
Homewood AE: Chiropractic and emotional disorders. In Schwartz HS (ed): Mental Health and Chiropractic, A Multidisciplinary Approach. New Hyde Park, NY, Sessions, 1973, pp 94-106.
Homewood AE:
The Neurodynamics of the Vertebral Subluxation, ed 3.
Place of publication not shown, published by author, 1981, pp 21, 250.
Hubbard JI:
The Peripheral Nervous System.
New York, Plenum, 1974.
Illis LS:
Regeneration in the central nervous system.
Lancet, 1:1035-1037, 1973.
Inglis BD, Fraser B, Penfold BR:
The scientific basis for chiropractic.
In Inglis BD, Fraser B, Penfold BR (eds): Chiropractic in New Zealand.
Davenport, IA, Palmer College of Chiropractic, 1979, pp 200-226.
Iversen LL;
The chemistry of the brain.
Scientific American, 24(3):134, 1979.
Jacobson E:
Tension in Medicine.
Springfield, IL, Charles C. Thomas, 1967.
Jacquet P:
Bioenergetic analysis and chiropractic: Theoretical considerations and practical application.
Annals of the Swiss Chiropractors' Association, 7:208-218, 1981.
Janse J:
Principles and Practice of Chiropractic.
Lombard, IL, National College of Chiropractic, 1976, pp 47-48.
Jaskoviak PA, Schafer RC:
Applied Physiotherapy: Practical Applications Within Clinical Chiropractic.
Arlington, VA, American Chiropractic Association, 1986, pp 3, 16.
Jewett DL, Rayner MD:
Basic Concepts of Neuronal Function.
Boston, Little, Brown, 1983.
Kandel ER:
Small systems of neurons.
Scientific American, 24(3):66, 1979.
Katz B:
Nerve, Muscle, and Synapse.
New York, McGraw-Hill, 1966.
Kimber DC, Gray CE, Stackpole CE:
Textbook of Anatomy and Physiology, ed 11.
New York, Macmillan, pp 186-268.
Kimmel EH:
Psychophysiological aspects of the subluxation.
In Schwartz HS (ed): Mental Health and Chiropractic, A Multidisciplinary Approach.
New Hyde Park, NY, Sessions, 1973, pp 135-147.
Kuffler SW, Nichols JG:
From Neuron to Brain.
Sunderland, MA, Sinauer Associates, 1976.
Kupfermann I:
Modulatory actions of neurotransmitters.
Annual Review of Neuroscience, 2:447, 1979.
Kuntz A:
The Autonomic Nervous System.
Philadelphia, Lea & Febiger, 1953.
Leach RA:
The Chiropractic Theories, ed 2.
Baltimore, Williams & Wilkins, 1986.
Levi-Montalcini R, Calissano P:
The nerve growth factor.
Scientific American, 240(6):68, 1979.
Lovgren PW.
Principles of treatment in chiropractic.
Scandinavian Journal of Rehabilitation Medicine, 11(4):157-162, 1979.
Majno G:
The Healing Hand.
Cambridge, MA, Harvard University Press, 1975.
McLennon H:
Synaptic Transmission.
Philadelphia, W.B. Saunders, 1963.
McNaught AB, Callander R:
Illustrated Physiology, ed 4.
New York, Churchill Livingstone, 1983, pp 20-21, 204-209, 211, 213, 219.
Mears D:
Mental disease and cervical spine distortion.
ACA Journal of Chiropractic, 2(9), September 1965.
Nelson WA:
Function vs structure.
ACA Journal of Chiropractic, IV:7-8, February 1970.
Noback CR, Demarest RJ:
The Nervous System: Introduction and Review.
New York, McGraw-Hill, 1972.
Ochs S:
Axoplasmic transport a basis for neural pathology.
In Dyck PJ, Thomas PK, Lambert EH (eds): Peripheral Neuropathy.
Philadelphia, W.B. Saunders, Chapter 12.
Offner F:
The excitable membrane.
Biophysical Journal, 12:1583-1629, 1972.
Olsson Y:
Vascular permeability in the peripheral nervous system.
In Dyck PJ, Thomas PK, Lambert EH (eds): Peripheral Neuropathy.
Philadelphia, W.B. Saunders, 1975, Chapter 10.
Palmer DD:
Textbook of the Science, Art and Philosophy of Chiropractic.
Portland, OR, Portland Printing House, 1910.
Pappas GD:
Junctions between cells.
In Weissman G, Claiborne R (eds): Cell Membrane: Biochemistry, Cell Biology, and Pathology.
New York, HP Publications, 1975, Chapter 9.
Parke WW:
Applied anatomy of the spine.
In Rothman RH, Simeone FA (eds): The Spine.
Philadelphia, W.B. Saunders, 1975, vol I, pp 28-31.
Patton HD, et al:
Introduction to Basic Neurology.
Philadelphia, W.B. Saunders, 1976.
Peele TL:
The Neuroanatomic Basis for Clinical Neurology, ed 2.
New York, McGraw-Hill, 1961.
Pinkenburg C:
A basis for the theory of manipulative medicine.
Journal of Manipulative and Physiological Therapeutics, 3(2):81-85, June 1980.
Pottenger FM:
Symptoms of Visceral Disease, ed 6. St. Louis, C.V.
Mosby, 1949, pp 74-76.
Prichard JW: Nerve.
In Albright JA, Brand RA (eds): The Scientific Basis of Orthopaedics.
New York, Appleton-Century-Crofts, 1979, pp 385-408.
Quigley WH:
Case Histories of Mental Illness Under Chiropractic Care. Davenport, IA,
Clear View Sanitarium, 1954, 1968.
Quigley WH:
Physiological psychology of chiropractic in mental disorders.
In Schwartz HS (ed): Mental Health and Chiropractic, A Multidisciplinary Approach.
New Hyde Park, NY, Sessions, 1973, pp 107-119.
Rupert RL, Wagnon R, Thompson P, Moh TE:
Chiropractic adjustments: Results of a controlled clinical trial in Egypt.
ICA Review, pp 58-60, Winter 1985.
Schwartz HS (ed):
Mental Health and Chiropractic, A Multidisciplinary Approach.
New Hyde Park, NY, Sessions, 1973.
Schwartz HS:
Foundations of chiropractic psychosomatics.
Journal of the National Chiropractic Association, June 1949.
Schwartz HS:
Preliminary analysis of 350 mental patients records treated by chiropractors.
Journal of the National Chiropractic Association, November 1949.
Shepherd GM:
Microcircuits in the nervous system.
Scientific American, 238(2): 92, 1978.
Singer M:
The influence of the nervous system on the regeneration of body parts.
Osteopathic Annals, 3(7):217-219, 1975
Singer SJ:
A Short History of Anatomy and Physiology from the Greeks to Harvey.
New York, Dover Press, 1957.
Speransky AD:
A Basis for the Theory of Medicine; translated by C.P. Dutt.
New York, International Publishers, 1943.
Stedman's Medical Dictionary, ed 24.
Baltimore, Williams & Wilkins, 1984.
Stevens CF:
The neuron.
Scientific American, 24(3):54, 1979.
Strang VV:
Essential Principles of Chiropractic. Davenport, IA,
Palmer College of Chiropractic, 1984, pp 48-58.
Sulzer JL:
Chiropractic healing as psychotherapy.
In Schwartz HS (ed): Mental Health and Chiropractic, A Multidisciplinary Approach.
New Hyde Park, NY, Sessions, 1973, pp 155-160.
Thomas CL (ed):
Taber's Cyclopedic Medical Dictionary, ed 14.
Philadelphia, F.A. Davis, 1981.
Thorpe RG:
Musculoskeletal factors in chronic disease.
Osteopathic Annals, 3(1): 7-10, 1975.
Vear HC:
An assessment of chiropractic scope of practice.
In Coyle BA, Martinez DP: Proceedings of the Second Conference on Current Topics in Chiropractic: Reviews of the Literature, May 4-5, 1985.
Sunnyvale, CA, Palmer College of Chiropractic West, 1985, p C1-2.
Verner JR:
The Science and Logic of Chiropractic, ed 4. Englewood, NJ,
published by author, 1950.
Vernon HT, Dhami MSI, Howley TP, Annett R:
Spinal manipulation and beta-endorphin: A controlled study of the effect of a spinal manipulation on plasma beta-endorphin levels in normal males.
Journal of Manipulative and Physiological Therapeutics, 9:115-123, 1986.
Wayne KM:
Disturbances of the nervous system as related to chiropractic.
Journal of the Canadian Chiropractic Association, 13(2):20-23, 1969.
Werner JK:
Trophic influence of nerves on the development and maintenance of sensory receptors.
American Journal of Physical Medicine, 53(3):127-142, 1974.
Weiant CW:
Rational Bacteriology. New York, New York
Institute of Chiropractic, 1953.
Zhigalina LI:
The role of nonspecific stimulation of the nervous system in certain immunological reactions.
In Gordienko AN (ed): Control of Immunogenesis by the Nervous System.
Washington, DC, US Department of Commerce, 1958, pp 110-124.