

Conjugated Linoleic Acid: A Review This section is compiled by Frank M. Painter, D.C.
Send all comments or additions to: Frankp@chiro.org


FROM: Alternative Medicine Review 2001 (Aug); 6 (4): 367–382 ~ FULL TEXT
Gregory S. Kelly, NDConjugated linoleic acid (CLA) refers to a group of positional and geometric isomers of the omega-6 essential fatty acid linoleic acid (cis-9, cis-12, octadecadienoic acid). In humans evidence is currently ambiguous as to whether CLA supplementation has a significant effect on body composition. Despite favorable changes in lipid levels in animal models, a beneficial effect in humans has not yet been established. While some of the changes reported are consistent with an improved lipid profile, declines in HDL and increases in lipoprotein (a) have also been observed in some subjects. Available evidence suggests CLA supplementation has no impact on immune system performance in healthy subjects.
Effect of CLA on Insulin Resistance
While CLA has received a great deal of attention as a supplement that might favorably modify body composition, its potentially adverse impact on insulin resistance has not received equal attention. If CLA has the effect on insulin levels and activity in human patient populations that it has had in some experimental animal studies, such action would be potentially undesirable.
In Vivo Research in Animals
In several animal studies, dietary CLA has apparently given rise to increased insulin levels. [16, 17, 30, 33] In AKR/J mice (a strain susceptible to dietary obesity), adding CLA as one percent of dietary calories resulted in a nearly two-fold increase in plasma insulin levels. In these mice there was also a trend toward higher blood glucose levels. [16] The combination of both higher circulating insulin and glucose levels suggests impairment in the ability of insulin to dispose of glucose in tissues, a finding consistent with the development of an insulin resistant state. Even though these mice had a reduction in adipose tissue weight, the development of an insulin resistant state would not be construed as a beneficial metabolic change.
In female mice fed standard semi-purified diets (10% of total energy as fat) with or without CLA (1%), fat mass decrease in these animals was attributed to apoptosis. Tumor necrosis factor-alpha (TNF-a) levels increased 12-fold in isolated adipocytes from CLA-fed mice compared with control mice. [19] TNF-a increases have been reported to be associated with insulin resistance and abdominal obesity in humans. [31] In this study's mice, the decrease in fat mass subsequent to CLA supplementation resulted in a state resembling lipoatrophic diabetes; i.e., ablation of brown adipose tissue, a marked reduction of white adipose tissue, marked hepatomegaly, and marked insulin resistance. [19] Other studies on mice fed CLA have also found increases in circulating insulin levels, in liver weight, [17, 30] and in TNF-a. [32]
The effect of CLA administration on insulin levels, glucose tolerance, and glucose homeostasis was investigated in male Zucker diabetic rats (an animal model of type 2 diabetes). When CLA-fed rats were compared with lard-fed rats, a reduction in insulin levels was seen in the CLA group; however, the CLA-fed rats remained markedly hyperinsulinemic. An improvement in glucose disposal and a more rapid return to baseline glucose levels subsequent to a glucose infusion was also observed in the CLA-fed rats. [33] While CLA feeding was superior to lard feeding in these genetically susceptible rats, it is important to note that these rats developed a substantial degree of insulin resistance as indicated by insulin levels approximately 10.5 times higher than levels found in lean controls. [33]
Feeding CLA to adult female pigs was reported to increase fasting insulin levels. One group of female pigs was fed a diet containing one-percent CLA. These animals were compared with another group fed an isoenergetically identical diet containing no CLA. After six weeks, no differences in body weights were found between the two groups; however, the CLA-treated pigs exhibited a 37-percent higher concentration of fasting serum insulin. [34]
In Vivo Human Research
Insulin levels have been monitored only in one human trial of CLA supplementation. Among women (ages 20-41 years) who were confined to a metabolic suite for a 94-day period, 3 g/day CLA resulted in an insignificant trend toward increased mean insulin levels toward the end of the supplementation period. CLA supplementation did not change blood glucose levels during the treatment period. [29]
Effect of CLA on Cardiovascular Disease
Although CLA has received attention suggesting it might benefit cardiovascular health, [35] there is currently no evidence to support the hypothesis that CLA protects against atherogenesis in humans. Even in animal studies, adding CLA to a diet that would otherwise produce atherosclerosis resulted in mixed results depending on the type of animal and, even within animals of the same species, the genetic strain being studied. Similarly, the ability of CLA administration to positively modify lipid levels has not occurred in all animal models. The one available human trial did not demonstrate clinically significant improvement in lipid levels and no efforts were made to directly monitor atherosclerosis.
In Vivo Research in Animals
Atherosclerosis can be induced in New Zealand white rabbits fed a semi-purified diet containing 0.1-0.2-percent cholesterol. Supplementing this diet with CLA at a level as low as 0.1 percent can inhibit atherosclerosis. Even among the rabbits with established atherosclerosis, adding CLA to the semi-purified diet seems to be beneficial. Adding one-percent CLA to the diet for 90 days resulted in an average regression of established atherosclerosis of 30 percent. [36]
CLA supplementation in conjunction with an atherosclerotic diet fed to rabbits showed similar beneficial effect in retarding atherosclerosis in a second study. The diet contained 14-percent fat and 0.1-percent cholesterol. CLA as 0.5 percent of the diet was fed to 50 percent of the rabbits. Lipid levels among the CLA-supplemented rabbits were considered to be more favorable, consisting of lower triglycerides and low density lipoprotein (LDL) cholesterol levels. Examination of the aortas of CLA-fed rabbits showed less atherosclerosis. [23]
Hamsters fed CLA-containing diets (0.06-, 0.11-, or 1.1% CLA) collectively had significantly reduced levels of plasma total cholesterol, very low density lipoprotein (VLDL) and LDL cholesterol, and triglycerides. No effects were seen on high density lipoprotein (HDL) cholesterol levels. Plasma tocopherol/total cholesterol ratios determined from plasma pools for CLA supplemented animals were significantly increased (86% greater in animals fed 1.1% of diet as CLA). Since no additional tocopherol was contained in the diets of the animals fed CLA, these results suggest CLA had a tocopherol-sparing effect. Morphological assessment revealed less early atherosclerosis in the CLA-supplemented animals. [37] Other studies have shown similar favorable modifications in lipid profiles subsequent to CLA-feeding in hamsters. [25, 39]
However, CLA does not appear to produce identical results in all animal models of atherosclerosis. Serum of adult female CLA-treated pigs (1% of diet) showed a trend toward increased levels of triglycerides and VLDL and LDL cholesterol, without distinct changes in the HDL fraction. The LDL:HDL cholesterol ratio was significantly increased by feeding CLA to these animals. [34]
In C57BL/6 mice, CLA (0.5% of the diet) was substituted for linoleic acid in the diet. This strain of mice is genetically susceptible to the development of fatty streaks in the intima of the aortic sinus when placed on a diet similar to the control diet used in the study. In these mice, CLA addition to the diet resulted in a lipid profile considered less atherogenic (lower triglycerides, and a higher serum HDL:total cholesterol ratio). Despite the more favorable serum lipid profile, CLA supplemented animals demonstrated a greater degree of development of fatty streaks in aortic tissue than did mice fed the control diet. [38] Since in this study the lipid profile was positively modified and yet atherosclerosis was actually promoted, it might not be prudent to assume that an improvement in lipid profiles subsequent to CLA-feeding in other animal models actually means the risk of atherosclerosis has been lowered.
In Vivo Human Research
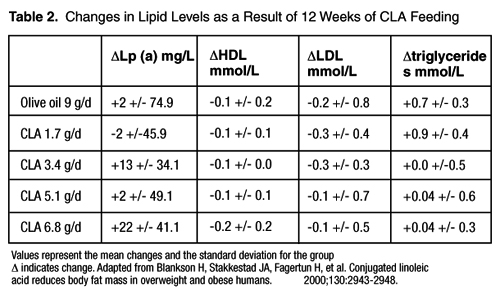
In a study of human subjects with BMIs between 25-35 kg/m2, statistically significant reductions in LDL, HDL, and total cholesterol were found in all groups receiving CLA; however, these changes were not large enough to be clinically significant. An increase in lipoprotein (a) was found in the groups receiving 3.4 grams or more of CLA per day. The average increase during the 12-week trial in the group receiving 3.4 grams per day of CLA was 13 +/- 34.1 mg/L. The average increase during the 12-week trial in the group receiving 6.8 grams per day of CLA was 22 +/- 41.1 mg/L. Data are summarized in Table 2. [27]
Effect of CLA on Cancer
The anticancer potential of CLA has received a great deal of research attention in both in vitro and animal models.
In Vitro Research
Several studies have demonstrated in vitro anticancer activity for CLA. CLA inhibited the proliferation of human hepatoma cell lines. [39] In vitro, CLA has shown inhibitory effects on lung adenocarcinoma cell lines (A-427, SK-LU-1, A549) and one human glioblastoma cell line. Unlike CLA, linoleic acid had no ability to inhibit growth of these cancer cell lines. [40] CLA inhibited proliferation of estrogen receptor-positive MCF-7 cells; however, it was ineffective in preventing proliferation of estrogen receptor-negative MDA-MB-231 cells. [41]
While CLA in the form of free fatty acids exerts an in vitro antiproliferative effect, milk fat enriched with CLA appears to have even greater in vitro activity. Bovine milk fat enriched with CLA was more effective in inhibiting human MCF-7 breast cancer cells than were isolated CLA isomers. Incubation of cells with bovine milk fat enriched with CLA decreased viable cell numbers by up to 90 percent. Incubation with a mixture of CLA isomers or with the c-9, t-11 CLA isomer resulted in a 60-percent decrease. Incubation with the t-10, c-12 CLA isomer caused only a 15-percent decrease in cell numbers under similar conditions. In contrast to the results obtained with the various forms of CLA, incubation with linoleic acid resulted in a 25-percent increase in cell proliferation. [42]
In Vivo Animal Research
In animal experiments, CLA has demonstrated anticancer activity and antimetastatic activity. Dietary CLA has anticancer activity in animal models of prostate [43] and colon [44] cancer; however, the majority of animal cancer research has studied the effects of CLA administration on mammary cancer.
A DU-145 human prostatic carcinoma cell line was injected into severe combined immunodeficient (SCID) mice. The mice were divided into three groups and tumor growth and metastasis were observed. One group was placed on a standard diet. The two remaining groups had a standard diet supplemented with either linoleic acid or CLA at one percent of the diet. Supplementation was started two weeks prior to subcutaneous inoculation with the DU-145 cells and was continued for a period of 14 weeks. The mice receiving CLA had smaller local tumors and a reduction in lung metastasis compared to the animals on both the standard diet and the linoleic acid supplemented diet. [43]
F344 rats were exposed to the carcinogen 2-amino-3-methylimidazol[4,5-f] quinolone in order to induce colon carcinogenesis. Controls and CLA treated rats (at 0.5% of the diet) were exposed to the carcinogen during weeks 3 and 4 of the study period. After week 16 the rats were killed in order to quantify aberrant crypt foci (ACF). While CLA administration had no impact on the size of ACF, the number of ACF was significantly reduced in the CLA group (1.1 +/- 1.3) when compared with the controls (4.3 +/- 2.4). [44]
The timing during the life cycle when CLA is added to the diet appears to exert a profound effect on protection against carcinogens that can initiate mammary tumor formation in mice and rats. It appears the critical time period for dietary exposure to CLA might be prior to mammary gland maturation. Several studies have found that dietary exposure to CLA between the time of weaning and mammary gland maturation has a protective effect against development of tumors due to exposure to carcinogens.
Feeding female rats a diet containing one-percent CLA from weaning until 50 days of age (corresponding with mammary gland maturity) resulted in a significant degree of protection against cancer formation due to dimethylbenz[a]anthracene (DMBA) exposure. In this study, no added protection against cancer formation was gained when the animals fed CLA from weaning to day 50 were maintained on a diet containing CLA after carcinogen exposure. [45]
In another study, female rats received no dietary CLA from weaning through 50 days of age (corresponding to mammary gland maturity). At day 50, rats were exposed to DMBA and CLA at one percent of the diet was initiated in all rats exposed to the carcinogen. CLA supplementation of the diet was discontinued at four weeks or eight weeks post-carcinogen exposure in some rats, and was continued until the end of the research experiment in other rats (20 weeks). Significant cancer protection was found only in the rats receiving CLA for the entire 20 weeks. As soon as CLA feeding was discontinued, protection against carcinogen-induced cancer formation was lost. [46]
When female rats were fed a diet containing one-percent CLA between early post-weaning and a period analogous to puberty (from 21 to 42 days of age), tumor formation as a result of methylnitrosourea (MNU) administration at 56 days of age was substantially reduced. The rats required no additional CLA supplementation during or after MNU exposure to maintain protection against mammary tumor initiation. In rats not exposed to dietary CLA during the post-weaning to pubertal period, tumor formation subsequent to MNU exposure was maximally prevented only if CLA was continuously supplemented starting after the MNU exposure. If CLA was withdrawn at any point after the MNU exposure in the animals not provided CLA during the period when breast tissue was maturing, tumor inhibition appeared to be lost. [47]
Temporary addition of CLA to the diet subsequent to mammary gland maturation did not provide protection against transplanted metastatic murine mammary tumors in female Balb/c mice. Eight-week-old female mice were fed diets containing 0.1-, 0.3- or 0.9-percent CLA for two weeks prior to transplantation of tumor cells into the mammary gland. Dietary CLA did not impact mammary tumor latency and provided no protection against tumor incidence. [48] In a study of SCID mice, continuous CLA administration (1% of diet), beginning two weeks prior to inoculation of human breast adenocarcinoma cells and continuing throughout the study time period, inhibited local tumor growth and tumor metastasis to lungs, peripheral blood, and bone marrow. [49] These studies taken together suggest that in mice, similar to results reported in female rats, continuous CLA feeding protected against tumor formation.
Addition of CLA to the diet of mice has shown antimetastatic effects in a murine mammary cancer model. Mice were fed a diet consisting of 20-percent fat. Diets were supplemented with either no CLA or 0.1-, 0.5-, or 1.0-percent CLA. Latency, metastasis, and pulmonary tumor burden of transplantable murine mammary tumors were measured. Compared with mice fed a diet containing no CLA, the latency of tumors in mice receiving CLA was significantly increased. Metastasis, as indicated by pulmonary tumor burden, was decreased in mice receiving CLA in the diet. Tumor burden was decreased proportionately with the increasing concentrations of dietary CLA, suggesting a dose-dependent response. [50]
Experimental results suggest a partial selectivity of uptake and accumulation of the c-9, t-11 CLA isomer in the mammary gland of rats. Since c-9, t-11 CLA appears to have greater in vitro antiproliferative ability, [42] and since it accumulates to a greater degree in rat mammary tissue, it has been suggested this isomer might be the most important for observed anticancer effects. [14, 42]
It appears the dose response to CLA is maximized at a dietary concentration of approximately one percent, and no added protection against mammary carcinogenesis is gained by increased concentrations of dietary CLA. [52] It also appears CLA prevents mammary cancer development subsequent to carcinogen exposure irrespective of the quantity (from 2-20% of the diet) and type (whether linoleic acid containing vegetable oils, arachidonic acid containing lard, or a combination of both) of other fats found in the diet. [51, 52]

Return to OMEGA-3 FATTY ACIDS
Since 10-01-2001
{kind=link}
| Home Page | Visit Our Sponsors | Become a Sponsor |
Please read our DISCLAIMER |