

Immobilization Induces Changes
in Presynaptic Control of
Group Ia Afferents in Healthy HumansThis section is compiled by Frank M. Painter, D.C.
Send all comments or additions to: Frankp@chiro.org


FROM: J Physiol. 2008 (Sep 1); 586 (Pt 17): 4121–4135 ~ FULL TEXT
OPEN ACCESS Jesper Lundbye-Jensen and Jens Bo Nielsen
Department of Neuroscience and Pharmacology,
University of Copenhagen,
The Panum Institute 22.3, Blegdamsvej 3, DK-2200
Křbenhavn N, Denmark.
Neural plasticity occurs throughout adult life in response to maturation, use and disuse. Recent studies have documented that H-reflex amplitudes increase following a period of immobilization. To elucidate the mechanisms contributing to the increase in H-reflex size following immobilization we immobilized the left foot and ankle joint for 2 weeks in 12 able-bodied subjects. Disynaptic reciprocal inhibition of soleus (SOL) motoneurons and presynaptic control of SOL group Ia afferents was measured before and after the immobilization as well as following 2 weeks of recovery. Following immobilization, maximal voluntary plantar- and dorsiflexion torque (MVC) was significantly reduced and the maximal SOL H-reflex amplitude increased with no changes in the maximal compound motor response (M(max)). Decreased presynaptic inhibition of the Ia afferents probably contributed to the increase of the H-reflex size, since we observed a significant decrease in the long-latency depression of the SOL H-reflex evoked by peroneal nerve stimulation (D2 inhibition) and an increase in the size of the monosynaptic Ia facilitation of the SOL H-reflex evoked by femoral nerve stimulation. These two measures provide independent evidence of changes in presynaptic inhibition of SOL Ia afferents and taken together suggest that GABAergic presynaptic inhibition of the SOL Ia afferents is decreased following 2 weeks of immobilization. The depression of the SOL H-reflex when evoked at intervals shorter than 10 s (homosynaptic post-activation depression) also decreased following immobilization, suggesting that the activity-dependent regulation of transmitter release from the afferents was also affected by immobilization. We observed no significant changes in disynaptic reciprocal Ia inhibition. Two weeks after cast removal measurements returned to pre-immobilization levels. Together, these observations suggest that disuse causes plastic changes in spinal interneuronal circuitries responsible for presynaptic control of sensory input to the spinal cord. This may be of significance for the motor disabilities seen following immobilization as well as the development of spasticity following central motor lesions.
From the Full-Text Article:
Introduction
It is well established that motor skill training may induce use-dependent functional and structural plasticity within the central nervous system. Experience-driven central nervous plasticity has been extensively documented at a cortical level in relation to motor learning (Buonomano & Merzenich, 1998; Sanes & Donoghue, 2000) and there is growing evidence that the cortical plasticity accompanying motor skill learning is paralleled by changes in the properties of the spinal neuronal circuitries as demonstrated in monkey (Wolpaw et al. 1983a,b), rats (Chen & Wolpaw, 1995), in the spinal central pattern generator properties in cats (de Leon et al. 1998; Pearson, 2000; Barriere et al. 2008) and human subjects following skill training (Perez et al. 2005; Meunier et al. 2007; Wolpaw, 2007).
It is less well investigated how and to what extent immobilization is also accompanied by changes in transmission in spinal neuronal circuitries. Spinal cord plasticity can occur at numerous neuronal and synaptic sites and through a variety of mechanisms. In humans, spinal cord plasticity has mostly been inferred from modifications in the size of H-reflexes (Perez et al. 2005, 2007; Gruber et al. 2007; Meunier et al. 2007). The H-reflex provides a gross measure of motoneuron pool excitability and transmission across the synapses of Ia afferents and thus reflects both presynaptic mechanisms, postsynaptic inhibition and excitation, intrinsic motoneuronal mechanisms and descending inputs.
Recent studies have provided evidence that H-reflex amplitudes increase following a period of immobilization. In the rat, Anderson et al. (1999) reported increases in the H-reflex gain following 3 weeks of hindlimb unloading and in human, Clark et al. (2006b) similarly observed increased H-reflex amplitudes at rest following 4 weeks of lower limb suspension. We have recently demonstrated an increase in the excitability of H-reflex following a period of 1 week of wrist and hand immobilization (Lundbye-Jensen & Nielsen, 2008). In this study, the size of the evoked H-reflexes increased with no significant changes in corticospinal excitability estimated from motor evoked potentials (MEPs) evoked by transcranial magnetic stimulation. Although this finding does not provide evidence of the specific mechanisms involved, it led to the hypothesis that immobilization may be accompanied by changes in the amount of presynaptic inhibition of the Ia afferents or post-activation depression (homosynaptic depression). It has previously been demonstrated that a considerable part of the H-reflex amplitude modulation is explained by changes in presynaptic inhibition of the synapses of Ia afferents on spinal motoneurons (Fig. 1; Hultborn et al. 1987a,b; Meunier & Pierrot-Deseilligny, 1989; Nielsen & Kagamihara, 1993b; Faist et al. 1996).
Figure 1: Spinal mechanisms:
During the experiments,
EMG was obtained from SOL and TA muscles. Peripheral
nerve electrical stimulation was delivered to the
posterior tibial nerve (PTN), the common peroneal
nerve (CPN) and the femoral nerve (FN) in order to
investigate disynaptic reciprocal inhibition,
‘classical’ presynaptic inhibition and
post-activation depression.
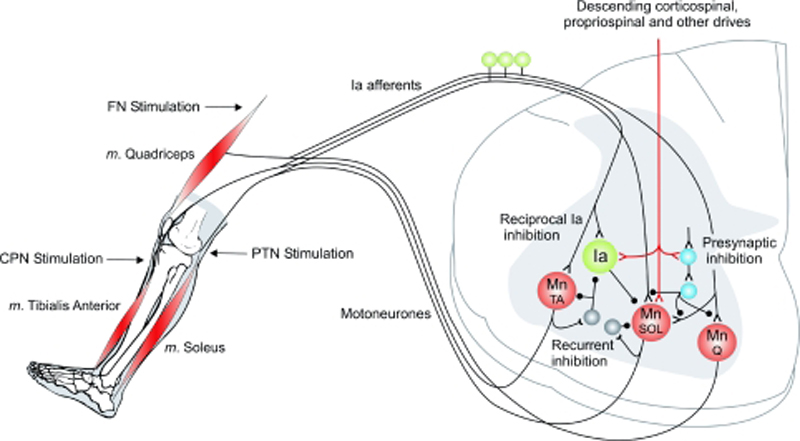
Presumably, gating of sensory inputs to spinal motoneurons is of functional importance in regulating the contribution of the stretch reflex circuitry to the ongoing motor activity (Nielsen & Sinkjaer, 2002; Rudomin, 2002). One advantage of regulating this contribution at the presynaptic level is that sensory inputs may be selectively gated, while allowing effective activation of the muscles by central commands (Rudomin, 2002). Furthermore, regulation of sensory information at the presynaptic level may permit conveyance of proprioceptive information to the cortex, which might contribute to adjusting supraspinal motor commands (Llewellyn et al. 1990; Morita et al. 1998). In line with this notion, both GABAergic presynaptic inhibition and post-activation depression have been demonstrated to increase following skill learning (Perez et al. 2005; Meunier et al. 2007), and it is possible that presynaptic control of sensory input changes during a period of immobilization. If this is the case it would be consistent with the findings of Manabe et al. (1989) who observed a disuse-induced enhancement of Ia synaptic transmission in spinal motoneurons of the rat.
The hypothesis that immobilization induces modifications of the transmission in the monosynaptic Ia pathway through changes in presynaptic control mechanisms was consequently tested in the present study.
We recorded the soleus H-reflexes before and after 2 weeks of ankle joint immobilization and following a corresponding period of recovery. Disynaptic reciprocal inhibition was evaluated through conditioning the SOL H-reflex by stimulation of the common peroneal nerve (CPN). To evaluate presynaptic inhibition of the soleus Ia afferents we measured the size of the femoral nerve monosynaptic Ia facilitation of the soleus H-reflex (Hultborn et al. 1987a,b) and the long-latency (D2) inhibition of the soleus H-reflex induced by CPN nerve stimulation (Mizuno et al. 1971). To assess post-activation (homosynaptic) depression we used the frequency-related depression of the Sol H-reflex (Crone & Nielsen, 1989b; Hultborn et al. 1996; Kohn et al. 1997).
Discussion
The primary purpose of the present study was to elucidate adaptations in spinal neuronal circuitries in relation to immobilization. The main behavioural finding was that 2 weeks of ankle joint immobilization leads to significant decreases of maximal voluntary strength. This was accompanied by an increase in the SOL H-reflex excitability in the immobilized leg as has previously been reported (Duchateau, 1995; Anderson et al. 1999; Clark et al. 2006b; Lundbye-Jensen & Nielsen, 2008). Based on the applied conditioning–test protocols of the SOL H-reflex, it appears that the observed increase of H-reflex excitability at least in part relates to changes in presynaptic control of group Ia afferents following immobilization whereas disynaptic reciprocal inhibition was unaffected by the immobilization.
MVC and twitch contractions
In the present study, immobilization led to a 14% reduction of plantar flexion MVC and a 23% reduction of dorsiflexion MVC. These changes are in agreement with the findings in previous studies reporting 17% and 19% decreases in triceps surae MVC following 2 weeks (Gondin et al. 2004) and 3 weeks (Davies et al. 1987) of immobilization, respectively. Depending on the duration of the immobilization period, the observed decrease in voluntary strength has been attributed partly to changes in neural activation and partly to changes in muscle contractile properties (White et al. 1984; Davies et al. 1987; Duchateau & Hainaut, 1987; Duchateau, 1995; Seki et al. 2001a,b; Gondin et al. 2004; Clark et al. 2006a,b). Several studies have found decreases in voluntary activation following immobilization (Vandenborne et al. 1998; Kawakami et al. 2001; Gondin et al. 2004). Seki et al. (2001a) reported a restriction of motoneurone firing to lower rates following immobilization and reduced maximal motoneuronal firing rates following only 1 week of immobilization (Seki et al. 2001a, 2007). Judged from the plantar flexor twitch measures used in the present study, it is not unlikely that changes in muscle contractile properties may have contributed to the reduced voluntary strength observed following immobilization. The finding of increased twitch contraction torque (TT) and the tendency toward prolonged half-relaxation time (HRT) following immobilization is indeed in agreement with previous studies on short-term immobilization (White et al. 1984; Davies et al. 1987; Seki et al. 2001b; Gondin et al. 2004). The trend towards increased HRT has previously been suggested to relate to reduction in the reuptake of Ca2+ by the sarcoplasmatic reticulum following immobilization (Thom et al. 2001). Although immobilization was accompanied by changes in twitch contraction parameters in the present study, the exact origin of these alterations remains to be determined.
Interestingly, immobilization was accompanied by a larger reduction of the MVC for dorsiflexion compared to plantar flexion. One possible reason for this could be differences in muscle characteristics, fibre-type distribution and myosin heavy chain (MHC) expression demonstrating different sensitivities to inactivity. Secondly, ankle joint immobilization, while restricting TA and SOL contractions, does not completely immobilize the biarticular gastrocnemius muscle. Although twitch contraction measures changed for triceps surae as a whole, the fact that contraction of gastrocnemius was possible during the immobilization may have counteracted loss of plantar flexion MVC following immobilization. In line with this, Gondin et al. (2004) found SOL, but not gastrocnemius, EMG activity to be significantly reduced following ankle joint immobilization.
Increased Hmax/Mmax ratio
In the present study we observed an increase of the Hmax/Mmax ratio at rest following immobilization. It has previously been reported that the Hmax/Mmax ratio is increased following a period of immobilization in man (Duchateau, 1995; Clark et al. 2006b; Lundbye-Jensen & Nielsen, 2008) and hindlimb suspension in the rat (Anderson et al. 1999) and that the gain (Hslope/Mslope ratio) of the H-reflex is increased (Lundbye-Jensen & Nielsen, 2008).
Modulation of the H-reflex amplitude can result from a number of factors, including changes in GABAergic presynaptic inhibition of the Ia afferents, variation in the amount of Ia neurotransmitter release, and changes in the excitability of the motoneurones due to changes in either the membrane potential arising from excitatory or inhibitory input (postsynaptic) or the intrinsic properties of the neurones. Cormery et al. (2005) recently observed changes in the intrinsic properties of rat motoneurones following 2 weeks of hindlimb unloading. This was signified by increased rheobase, decreased spike amplitudes and deceased membrane time constants. Some properties of ‘slow’ motoneurones resembled those of ‘fast’ motoneurones following hindlimb unloading (Cormery et al. 2005). This finding indicates that the motoneurones are not more excitable or can be more easily activated. However, a selective change in the properties of the ‘slow’ motoneurones means that the changes observed in H-reflex following immobilization in the present study could be due to changes in the input–output relation or ‘recruitment gain’ of the motoneurone pool following immobilization (Kernell & Hultborn, 1990; Hultborn et al. 2004).
In a recent study, we observed increased H-reflex amplitudes following 1 week of wrist immobilization with no significant changes in corticospinal excitability estimated from transcranial magnetic stimulation measures (Lundbye-Jensen & Nielsen, 2008). Although many mechanisms may potentially be involved in this phenomenon, we hypothesized, based on this finding, that presynaptic control of Ia afferent input to the spinal cord could change (decrease) following a period of immobilization. If so, this would be in agreement with Manabe et al. (1989) who observed a disuse-induced enhancement of monosynaptic Ia afferent transmission (EPSPs) to rat spinal motoneurones following conduction block with tetrodotoxin and nerve section (Manabe et al. 1989).
Presynaptic inhibition of Ia afferents
It has previously been demonstrated that part of the H-reflex modulation during voluntary movement may be explained by changes in presynaptic inhibition of the synapses of Ia afferents on spinal motoneurones (Hultborn et al. 1987a,b; Meunier & Pierrot-Deseilligny, 1989; Nielsen & Kagamihara, 1993b; Faist et al. 1996; Perez et al. 2005). Presumably, gating of sensory input to spinal motoneurones is of functional importance in regulating the contribution of the stretch reflex circuitry to the ongoing motor activity (Rudomin & Schmidt, 1999; Nielsen & Sinkjaer, 2002). One advantage of regulating this contribution at the presynaptic level is that sensory inputs may be selectively gated while allowing effective activation of the muscles by central commands (Rudomin, 2002). Furthermore, regulation of sensory information at the presynaptic level may permit conveyance of proprioceptive information to the cortex which might contribute to adjusting of supraspinal motor commands (Llewellyn et al. 1990; Morita et al. 1998). There is good evidence from previous studies that changes in presynaptic inhibition of the synapses between sensory afferents and motoneurones is fundamental in the adaptation of the reflex circuitry during motor learning (Kandel et al. 2000; Wolpaw, 2007). Recently, Perez et al. (2005) demonstrated that presynaptic inhibition of Ia afferents from SOL was increased following a period of visuomotor learning. However, it has not to our knowledge been investigated whether presynaptic inhibition changes following a period of immobilization.
Presynaptic inhibition consists of a GABA-mediated depolarization of the terminals of primary afferents, which results in a decreased transmitter release (Schmidt, 1971; Rudomin, 1990). In animal experiments, the most direct demonstration of presynaptic inhibition of Ia afferents (see Eccles, 1964) is the depression of the monosynaptic EPSP evoked in motoneurones by a constant stimulation of Ia afferents without a change in the motoneurone membrane potential and conductance (Frank & Fuortes, 1957). In human studies, information about presynaptic inhibition can only be obtained indirectly.
In the present study, we observed significant increases in the heteronymous facilitation of the SOL H-reflex evoked by femoral nerve stimulation (Hultborn et al. 1987a) and a reduction in the amount of D2 inhibition. These two measures provide independent evidence of changes in presynaptic inhibition of SOL Ia afferents and taken together with the observation of unchanged disynaptic reciprocal inhibition these findings strongly suggest that presynaptic GABAergic inhibition of the SOL Ia afferents is reduced following 2 weeks of ankle joint immobilization.
The early facilitation of the SOL H-reflex evoked by femoral nerve stimulation provides indirect information about the heteronymous Ia EPSPs evoked in the motoneurone pool (Hultborn et al. 1987a). It is essential for the method that the investigated facilitation of the SOL H-reflex evoked by FN stimulation is monosynaptic in origin, as a change in facilitation may otherwise be caused by postsynaptic changes at an interneuronal level, meaning that changes in facilitation of the test reflex cannot be ascribed unequivocally to changes in the size of the conditioning Ia EPSP. This is why in the present study the onset of facilitation was examined in detail with conditioning–test intervals in steps of 0.2 ms. Considering the homonymous D2 inhibition, PAD interneurones mediating presynaptic inhibition of the Ia afferent volley of the test reflex are activated by a conditioning group I volley. Although the D1 and D2 inhibition (Mizuno et al. 1971) is probably contaminated by other effects (Meunier & Pierrot-Deseilligny, 1998), the size of these depressions can be assessed to estimate the excitability of PAD interneurones mediating presynaptic inhibition. Following immobilization, D2 inhibition decreased significantly, which also indicates that presynaptic inhibition is decreased. Decreased inhibition could result from decreased excitability of PAD interneurones or occlusion following activation by another excitatory input (Faist et al. 1996). However, in the lower limb, Iles (1996), using very weak stimuli, has shown that a cortically evoked decrease in CPN-induced inhibition is not due to occlusion in PAD pathways. Furthermore, in the present experiments, the decreased D2 inhibition was associated with increased heteronymous facilitation, which cannot be due to an occlusion in PAD pathways (see Hultborn et al. 1987b).
As previously mentioned, it may be argued that an increased heteronymous Ia facilitation could be observed as a consequence of skewed input to or effects within the motoneurone pool. This could potentially affect the recruitment gain by increasing the slope of the input–output relation of the H-reflex (Kernell & Hultborn, 1990; Nielsen & Kagamihara, 1993b). This would be in agreement with the findings of Cormery et al. (2005). However, if recruitment gain had been changed by a skewed distribution of effects within the motoneurone pool, this would be expected to enhance the effect of all synaptic input to the motoneurones thereby increasing D1 and D2 inhibition. In effect, the larger the recruitment gain the larger should be the D1 and D2 inhibition. The present finding of increased heteronymous Ia facilitation and decreased D2 inhibition following immobilization is in other words not consistent with a change in the recruitment gain. In conclusion, the observed changes are most probably caused by changes in presynaptic inhibition.
Post-activation depression
Post-activation depression also decreased following immobilization. Post-activation depression of the elicited reflex is caused by a frequency-dependent decrease in the probability of transmitter release from previously activated synapses. This depression of synaptic efficiency following previous synaptic activation of Ia afferents has been well characterized in both animal (Curtis & Eccles, 1960; Hultborn et al. 1996) and human studies (Crone & Nielsen, 1989a; Hultborn et al. 1996). We suggest that the long-term changes we observe in the frequency-related depression of the SOL H-reflex following immobilization reflect plastic changes at the level of the synapse between Ia afferents and SOL motoneurones causing an increase in the efficacy of the synapse. The molecular mechanisms underlying post-activation depression are not yet fully understood and subsequently we can only speculate about the possible mechanisms contributing to the modifications in the amount of transmitter released following immobilization.
It has, however, recently been reported by Meunier et al. (2007) that post-activation depression is subject to use-dependent plasticity. They reported that post-activation depression increased immediately following and 1 day after a single bout of skilful bicycle training. Contrary to the GABAergic presynaptic inhibition, there is so far no evidence for a descending control of post-activation depression, but some effects of, for instance, monoaminergic systems cannot be excluded. It is also not unlikely that structural modifications of the synapses between Ia afferents and SOL motoneurons have occurred during the 2 weeks of immobilization.
Post-activation depression is decreased in spastic patients (Faist et al. 1994; Nielsen et al. 1995) and Hultborn & Nielsen (1998) previously pointed out that the decreased post-activation depression in patients with cerebral or spinal lesions may (at least in part) be secondary to the disuse of motoneurons and Ia afferents accompanying the primary disorder. This notion is emphasized by the results of the present study and further supported by the findings of Gallego et al. (1979) that in the cat, prolonged disuse of the sensory fibres causes an increase in synaptic efficacy (EPSPs) of primary afferents in triceps surae motoneurons. Conversely, Meunier et al. (2007) demonstrated that motor training increased post-activation depression.
As already mentioned post-activation depression differs from the classical presynaptic inhibition transmitted through GABAergic axo-axonal synapses (see review by Hultborn & Nielsen, 1998). Even though both mechanisms contribute to the gating of sensory input to the central nervous system through modulation of synaptic transmitter release, the two mechanisms have distinct characteristics. Nevertheless, it is possible that an interaction between GABAergic presynaptic inhibition and post-activation depression contributes to the observed changes following immobilization. It has previously been suggested from animal studies (Enriquez-Denton et al. 2002) that reduced presynaptic inhibition also results in reduced post-activation depression and vice versa.
It is not possible from our data to make any conclusions regarding the exact mechanism(s) of the reduced presynaptic inhibition following immobilization, but some speculations may be made. There is good evidence from cat experiments that primary afferent synapses on ascending neurons and motoneurons are inhibited by different populations of interneurons (Jankowska & Padel, 1984). Indeed, the network responsible for controlling presynaptic inhibition of primary afferents seems in general to be organized to ensure selective control of afferent input to specific populations of both motoneurons and ascending neurons (Rudomin & Schmidt, 1999; Rudomin, 2002) and it is not impossible that changes in sensory input to the spinal interneurons that convey the presynaptic inhibition lead to plastic changes during a period of immobilization. According to afferent input and the functional requirements to the system, the CNS may increase or decrease the level of presynaptic inhibition of Ia afferents and thereby the central gain of the monosynaptic stretch reflex (Capaday & Stein, 1986; Hultborn et al. 1987a; Nielsen & Kagamihara, 1993b).
Based on the findings of the present study, it seems plausible that the decreased presynaptic inhibition occurred in response to the decreased motor activity accompanying limb immobilization. The changes in presynaptic inhibition may be a direct consequence of a reduced voluntary motor activity. It is, however, also possible that immobilization is accompanied by decreased proprioceptive input to the central nervous system. Reducing the amount of presynaptic inhibition of Ia afferents may in this way be a means of the central nervous system to increase the gain of the actual incoming afferent input. Since stimulation of low-threshold cutaneous mechanoreceptors by light brushing of both distal dorsal and plantar surfaces of the foot has also been demonstrated to decrease presynaptic inhibition (Iles, 1996), it may also be possible that the changes observed following immobilization occur in response to wearing the ankle cast. Measurements were, however, obtained after removal of the cast.
Functional and clinical perspectives
Although the present study involved limb immobilization in able-bodied subjects, the findings may also be of clinical relevance. This is especially the case in relation to neurological disorders leading to physical inactivity. It is noteworthy that the findings of increased H-reflexes, decreased GABAergic presynaptic inhibition and decreased post-activation depression following immobilization to some extent matches the findings of previous studies in spastic patients (Nielsen et al. 1993, 1995; Faist et al. 1994; Grey et al. 2008) and it is worth considering the effects of reduced physical activity in itself. As mentioned previously, it is possible that the decreased presynaptic inhibition and post-activation depression observed in patients with cerebral or spinal lesions may at least in part be a consequence of the disuse of motoneurons and Ia afferents.
Sensory feedback mechanisms through spinal reflex circuits help to ensure that the muscle activity is optimally adjusted to the immediate environment during movement. From a functional perspective it is interesting how changes in the functional properties of the central nervous system following a period of immobilization relate to functional motor control and further how subsequent training or rehabilitation affects these changes.
Acknowledgments
This work was supported by grants from The Danish Health Science Research Council, The Danish Ministry of Culture, The Novo Nordisk Foundation, The Carlsberg Foundation, The Elsass Foundation and the Danish Society of Multiple Sclerosis

Return to SUBLUXATION
Since 3-11-2013
| Home Page | Visit Our Sponsors | Become a Sponsor |
Please read our DISCLAIMER |