

DHA-rich Oil Modulates the Cerebral Haemodynamic Response
to Cognitive Tasks in Healthy Young Adults:
A Near IR Spectroscopy Pilot StudyThis section is compiled by Frank M. Painter, D.C.
Send all comments or additions to: Frankp@chiro.org


FROM: British J Nutrition 2012 (Apr); 107 (8): 1093–1098 ~ FULL TEXT
Philippa A. Jackson, Jonathon L. Reay, Andrew B. Scholey and David O. Kennedy
Brain, Performance and Nutrition Research Centre,
Northumbria University,
Newcastle upon Tyne,
NE1 8ST, UK.
philippa.jackson@northumbria.ac.uk
The impact of dietary omega-3 polyunsaturated fatty acids (PUFA) on behavioural outcomes has been widely researched; however, very little attention has been given to their impact on brain functioning in physiological terms. A total of twenty-two healthy adults took part in this double-blind, placebo-controlled study, wherein the cerebral haemodynamic effects of 12 weeks of daily dietary supplementation with either 1 g DHA-rich or 1 g EPA-rich fish oil (FO) or placebo (1 g olive oil) were assessed. Relative changes in the concentration of oxygenated hemoglobin (oxy-Hb) and deoxygenated Hb were assessed in the prefrontal cortex using near IR spectroscopy (NIRS) during the performance of four computerised cognitive tasks. Supplementation with DHA-rich FO, in comparison with placebo, resulted in a significant increase in the concentrations of oxy-Hb and total levels of Hb, indicative of increased cerebral blood flow (CBF), during the cognitive tasks. In comparison, no effect on CBF was observed following supplementation with EPA-rich FO, where concentration changes in the chromophores followed the same pattern as placebo. These encouraging pilot data warrant further application of NIRS in this area.
From the FULL TEXT Article:
Background
The impact of dietary omega-3 polyunsaturated fatty acids (PUFA) on human health has been increasingly examined in recent years. This unique class of fatty acids comprises the essential parent precursor fatty acid α-linolenic acid and principal metabolites include DHA and EPA. α-Linolenic acid cannot be manufactured de novo and must be supplied via dietary sources and is primarily found in seed oils such as flax and rapeseed. The more biologically relevant longer-chain fatty acids DHA and EPA can be synthesised from α-linolenic acid; however, this process is limited in humans. [1] A better dietary source of these fatty acids is therefore preformed DHA and EPA, which are abundant in oily fish. DHA is heavily concentrated in the mammalian brain and comprises 10–20 % of the fatty acids found in this organ. [2] DHA maintains the structural integrity of neuronal cell membranes and has an impact on numerous signalling pathways and neurotransmitter systems. [3, 4] EPA, although not a major component of the brain, could potentially influence brain function via the action of its many eicosanoid derivatives; regulators of key immune, endocrine and cardiovascular functions. [5, 6] A large body of research now exists describing the effects of dietary n-3 PUFA on behaviour; however, very little attention has been given to evaluating their impact on brain function in physiological terms, with only a limited number of preclinical and clinical trials to date addressing this issue. [7–9]
A viable approach to exploring the effects of dietary n-3 PUFA on brain function is by measuring cerebral haemodynamics using functional near IR spectroscopy (NIRS), a well-validated and non-invasive optical imaging technique. NIRS is based on the intrinsic optical absorption properties of oxygenated hemoglobin (oxy-Hb) and deoxygenated Hb (deoxy-Hb), and can be used as a measure of activation of neural tissue or simply as a measure of local cerebral blood flow (CBF) in the top layers of the cortex. Total-Hb (the sum of oxy- and deoxy-Hb) is closely related to cerebral blood volume, from which changes in regional CBF (rCBF) can be inferred. [10] Total-Hb also correlates well with rCBF changes as measured using the labelled water technique [11], although studies using rat brain models suggest that oxy-Hb is a better measure of rCBF than total-Hb. [12] NIRS has been successfully used to image activation in the frontal cortex (Fallgatter & Strik( [13, 14]) and other areas of the brain including the temporal, visual and parietal cortices (Schecklmann et al. [15]; Jasdzewski et al. [16]). Only a small number of trials published to date have utilised NIRS in pharmacological interventions (Kanamaru et al. [17]; Bonoczk et al. [18]); data from our own laboratory have recently demonstrated increased rCBF following administration of a vasodilator (resveratrol [19]) and decreased rCBF following a vasoconstrictor (caffeine [20]), supporting the application of NIRS in this area.
To date, research has primarly focused on the effects of DHA on brain function, given the high proportion of this fatty acid found in this organ. However, it is also possible that EPA could also have an impact on the cerebrovascular parameters, given that this fatty acid is readily incorporated into endothelial cell membranes elsewhere in the cardiovascular system [21] and has been shown to modulate local CBF in aged, stroke-prone rats. [22] Therefore, the objective of the present pilot trial was to investigate any treatment-related effects of 12 weeks of supplementation with DHA- and EPA-rich fish oil (FO) in healthy volunteers on the cerebral haemodynamic response to cognitive tasks in the prefrontal cortex.
Experimental methods
Participants
A total of twenty-two healthy adults took part in the study (nine male; mean age 21·96 years; mean BMI 23·04 kg/m2). All participants declared that they were in good health, non-smokers, free from prescription medication and social drugs, free from n-3 supplements and native English speakers. The participants also confirmed that they did not consume more than one portion of oily fish per week. At the time of testing, the participants were enrolled in a larger intervention trial assessing the cognitive and mood effects of 12 weeks of dietary supplementation with FO and were invited to take part in an extra testing session once they had completed their 12–week cognitive assessment. [23]
The present study was conducted according to the guidelines laid down in the Declaration of Helsinki, and all procedures involving participants were approved by the Northumbria University School of Life Sciences Ethics Committee. Written informed consent was obtained from all the participants.
Treatments
Before the start of the behavioural intervention study, a restricted randomisation list matching treatments to participant code numbers was generated by the computer. The participants were therefore randomly allocated to one of three treatment groups, DHA-rich FO, EPA-rich FO and placebo. The 1 g daily dose was provided by 2 × 500 mg capsules. Each active 500 mg capsule contained 497·5 mg of deodorised FO+2·5 mg mixed tocopherols. The total daily dose of n-3 PUFA provided by 2 × 500 mg capsules for the DHA-rich FO was 450 mg DHA+90 mg EPA (DHA 5:1 EPA), and for the EPA-rich FO, this was 300 mg EPA+200 mg DHA (EPA 3:2 DHA). The total daily dose for the placebo treatment was 1 g olive oil, again provided by 2 × 500 mg capsules. Treatment oils were purchased from EPAX AS (Aalesund, Norway) and encapsulated by Cardinal Health UK. All capsules used a brown bovine gelatine casing, and all treatments were packaged, labelled and randomised on site by a disinterested third party.
Near IR spectroscopy
The guiding principle of near IR spectroscopy (NIRS) is that the chromophores oxy-Hb and deoxy-Hb absorb light at different wavelengths. Therefore, by measuring the amount of light absorbance at a specific wavelength, the concentrations of oxy-Hb and deoxy-Hb can be calculated. [13] The third outcome produced by NIRS measurements – total Hb – is simply the sum of oxy-Hb and deoxy-Hb. In the present study, relative changes in the absorption of near IR light were measured using a 12–channel continuous-wave Oxymon system (Artinis Medical Systems B.V., Zetten, The Netherlands), described previously by Kennedy et al. [19] In this system, two nominal wavelengths of light (approximately 765 and 855 nm) are introduced through the skull via a laser emitter and measured, following transit through the upper surface of the cortex, by an optode placed at a preset distance from the light source (40 mm in this case). At this distance, the NIR signal is sensitive to haemodynamic changes within the top 2–3 mm of the cortex. [24] Given the experimental aims a simple two-channel configuration was used in this instance, with data collected from the areas of the frontal cortex corresponding to the International 10–20 system Fp1 and Fp2 EEG positions. NIRS data were time stamped by the researcher at the start and end of each task so that only NIRS data collected during the tasks were analysed. Recorded data were stored as a. TXT file and analysed using SPSS 16.0 (SPSS, Inc., Chicago, IL, USA).
Cognitive tasks
A total of four tasks were administered using the Computerised Mental Performance Assessment System, a software platform for the presentation of classic and bespoke computerised cognitive tasks. For accurate measurements, the NIRS optodes are required to make good contact with skin; therefore, the measures of haemodynamic response in the frontal cortex are most effective, given that the forehead is naturally free from hair. To this end, the tasks were selected on the basis that execution of the same or a similar task has previously been shown to activate the prefrontal cortex in brain imaging studies (Stroop task [25]; peg-and-ball task, 3–back task [26]; Wisconsin card sort task [27]).
The tasks were presented via a laptop computer in the same order as listed below. Progress through the battery of tasks was controlled by the participant, with brief instructions given onscreen before the start of each task. A 60 s rest period between each task was incorporated into the battery commencing as soon as the last response of the previous task had been made and ended with the presentation of the onscreen instructions for the next task. The cognitive data were not analysed.
The tasks were completed in the following order: Stroop task (5 min); peg-and-ball task (2·5 min); 3–back task (4 min); Wisconsin card sort task (3 min). Full descriptions of the Stroop, peg-and-ball and Wisconsin card sort tasks can be found in Kennedy et al. [28]. For the 3–back task, a continuous string of letters (upper and lower case; inter-stimulus interval of 2·5 s) was presented: seventy-eight letters in total with thirty-six target pairs. For each stimulus, the participants were instructed to indicate using a yes/no key press whether this was the same letter that appeared three letters previously.
Procedure
As part of a larger randomised, intervention trial investigating the effects of two types of FO on cognitive function and mood, the participants had been taking a daily 1 g supplement of either EPA-rich FO, DHA-rich FO or olive oil placebo for 12 weeks. Towards the end of the 12–week period, the participants were invited via email to take part in an extra cognitive testing session with concurrent NIRS data collection. In all cases, this assessment either took place on the same day as the final behavioural testing session or the following day (average time on treatment 88·59 (sem 3·9) d). Therefore, the final treatment dose was consumed 1 or 2 d before the NIRS testing session. Testing took place in the afternoon, and the participants were instructed to refrain from consuming food and drink except water for a minimum of 2 h before testing.
On arrival, the participants were seated in front of the laptop computer used for presenting the cognitive tasks. They were fitted with the NIRS headband and were instructed to sit quietly with their eyes closed until they were given further instruction by the researcher to start the cognitive tasks. Recording of NIRS data was initiated following 30 s of eyes-closed relaxation, and the participant was instructed to begin the tasks after a further 2 min of eyes-closed relaxation. This 2 min period was utilised as the NIRS resting baseline measurement. The participants completed each task with NIRS data collected throughout.
Treatment compliance

Table 1 The results for compliance as measured by capsule count were similar in all three treatment conditions (89 % DHA-rich FO, 87 % EPA-rich FO and 88 % placebo). As part of the behavioural intervention trial, the participants were required to give two venous blood samples, once before supplementation and again at the week 12 time point. The samples were collected and analysed as described by Jackson et al. (23); available data for the participants who completed the NIRS assessment are presented in Table 1.
Statistical methods
Changes in concentration (compared with resting baseline) of oxy-Hb and deoxy-Hb were averaged across the two channels (hemispheres) before analysis. Data collected during each task were averaged over the 2·5–5 min of task performance. The resultant data therefore represent the change in the concentration of each of the chromophores, plus their sum–total Hb–during each complete task period with respect to the pre-testing 2 min baseline measurement. As the duration of each task was substantially longer than the potential physiological oscillations that can cause a drift in shorter periods of the NIRS recording (e.g. heartbeat and respiration [29]), no adjustment was made to the data for this phenomenon.
The primary analysis of the averaged NIRS data was conducted by one-way ANOVA for each task with a priori planned comparisons of the data from each task being made between placebo and each treatment group (DHA-rich FO, EPA-rich FO) using t tests calculated with the mean squares error of the overall ANOVA. [30] To protect against an increased type I error rate, only those planned comparisons associated with a significant main effect of treatment on the ANOVA are reported.
Results
Oxygenated Hb
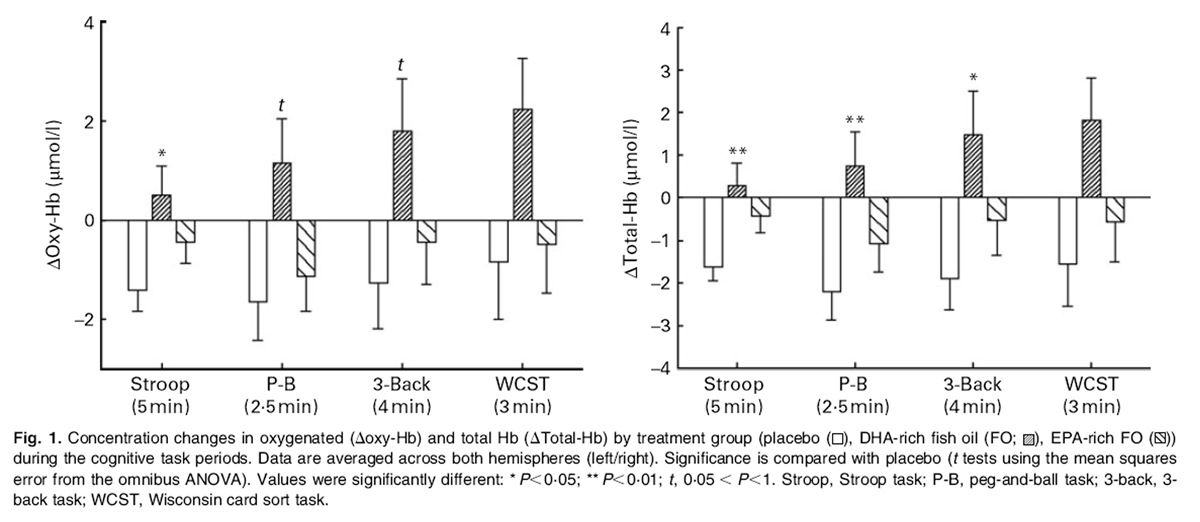
Figure 1 There was a significant main effect of treatment on oxygenated hemoglobin (oxy-Hb), while participants completed the Stroop task (F(2,19) = 3·72, P = 0·043). Reference to the planned comparisons showed a significant increase in the concentration of oxy-Hb following the DHA-rich FO during this task compared with placebo (P = 0·013). The trends for an effect of treatment were observed for the peg-and-ball and the 3–back tasks (F(2,19) = 3·35, P = 0·057; F(2,19) = 2·69, P = 0·093, respectively) (Figure 1).
Deoxygenated Hb
The ANOVA revealed no treatment-related effects on the concentrations of deoxy-Hb during any of the cognitive tasks.
Total Hb
The ANOVA showed that there was a significant effect of treatment on total-Hb during the Stroop, peg-and-ball and 3–back tasks (F(2,19) = 4·93, P = 0·019; F(2,19) = 4·21, P = 0·031; F(2,19) = 3·57, P = 0·048, respectively). Reference to the planned comparisons showed a significant increase in total-Hb in the DHA-rich FO treatment condition compared with placebo during all three of these tasks (Stroop (P < 0·006); peg-and-ball task (P < 0·009); 3–back task (P < 0·016) (Fig. 1).
Table 2 Planned comparisons did not reveal any significant difference in task-related concentration changes in any of the chromophores in the EPA-rich FO treatment condition, in comparison with placebo. Averaged data for each treatment group during the task periods are shown in Table 2.
Discussion
The present results reveal that supplementation with DHA-rich FO was associated with a significant increase in rCBF in the prefrontal cortex in response to cognitive task performance, in comparison with placebo, as indexed by changes in the concentration of oxy-Hb and total-Hb. Given that a similar pattern of results was not observed in participants who were administered EPA-rich FO, which also contained DHA, it would appear that the rCBF response to the tasks is only modulated following supplementation with DHA at a dose higher than 200 mg/d.
The pattern of concentration changes in oxy-Hb and total-Hb in the EPA-rich FO group did not differ from placebo, suggesting that the addition of dietary EPA at a dose of 300 mg/d for 12 weeks has no impact on cerebral haemodynamic response to cognitive tasks in this population. However, referring to the limited serum fatty acid data indicates that serum concentrations of EPA in this treatment group did not significantly increase following the 12–week intervention, despite a similarly high level of compliance with the treatment regimen being followed as in those administered the DHA-rich FO. Therefore, the impact of EPA supplementation on cerebrovascular response to cognitive tasks may require further exploration, and future investigations at doses higher than the present study would be advised. That being said, Sublette et al. [31] found significant correlations between cerebral metabolic rates of glucose (tempoparietal and frontal cortices, anterior cingulate) and peripheral concentrations of DHA and arachidonic acid, but not of EPA, suggesting instead that EPA may simply not have an impact on cortical activity, possibly due to, as these authors also predict, the very small amount of EPA found in the brain (approximately 1 % total fatty acids [2]).
The pattern of increased oxy-Hb and total-Hb in the DHA-rich FO treatment group, compared with placebo, is in agreement with recent data published by McNamara et al. [9] Using fMRI, these authors revealed that compared with placebo, DHA supplementation for 8 weeks resulted in increased activation of the dorsolateral prefrontal cortex during a sustained attention task in thirty-three schoolchildren (male, 8–10 years). It is worth noting that the NIRS parameters generated in the present study have been shown to closely correspond to the fMRI blood oxygen level-dependent signal [32], suggesting consistency of findings across these two paradigms. Interestingly, the authors also report null effects of the treatment on cognitive performance measures, despite the observed increase in cortical activity. Although the sample size in the present study was too small to permit an analysis of the behavioural data, performance measures analysed from the larger intervention trial described by Jackson et al. [23] from which the current sample was drawn revealed no effect of either active treatment (DHA-rich FO and EPA-rich FO) on cognitive performance overall.
The results from both studies suggest that the administration of DHA may improve the overall function and health of the brain, in terms of increased CBF, but that this is simply not manifested in a strong pattern of observable behavioural measures in young, healthy populations. A possibility is that improved function in terms of both haemodynamic response and behaviour may only be apparent in groups characterised by either natural or disease-related cerebrovascular insufficiency. Of course, it may also be the case that the length of the intervention in both our own study and the study described by McNamara et al. [9] was too short for any potential behavioural effects to be observed. However, longer intervention trials, in children at least, have reported similar null findings (Kirby et al. [33]; Osendarp et al. [34]).
Overall, the results of the present pilot study provide evidence of modulation of CBF parameters in healthy young adults following dietary supplementation with DHA-rich FO, and at a dose (1 g/d) that would be achievable by dietary intake of oily fish alone. Previously, studies that have aimed to evaluate the effect of n-3 PUFA supplementation on behavioural outcomes in healthy individuals have provided mixed results. However, the data from the present study suggest that an n-3 PUFA dietary supplement may indeed have an impact on cerebrovascular function. Given these interesting findings, further studies using NIRS and other imaging techniques to assess the causal relationship between n-3 PUFA intake and brain function are warranted.
Acknowledgements
The present study was completed as part of the PhD degree of P. A. J., which was funded by Ginsana SA, Switzerland, who also provided participant payments. Ginsana SA did not have any other input into the study design or analysis of the data. P. A. J. collected the data, and all authors contributed to the design, analysis and manuscript preparation. The authors declare no conflicts of interest.
References:
Burdge, GC & Calder, PC (2005)
Conversion of alpha-linolenic acid to longer-chain polyunsaturated fatty acids in human adults.
Reprod Nutr Dev 45, 581–597McNamara, RK & Carlson, SE (2006)
Role of omega-3 fatty acids in brain development and function: potential implications for the
pathogenesis and prevention of psychopathology.
Prostaglandins Leukot Essent Fatty Acids 75, 329–349Horrocks, LA & Farooqui, AA (2004)
Docosahexaenoic acid in the diet: its importance in maintenance and restoration of neural membrane function.
Prostaglandins Leukot Essent Fatty Acids 70, 361–372Innis, SM (2007)
Dietary (n-3) fatty acids and brain development.
J Nutr 137, 855–859 | PubMedPiomelli, D (1994)
Eicosanoids in synaptic transmission.
Crit Rev Neurobiol 8, 65–83 | PubMedCalder, PC (2006)
n-3 Polyunsaturated fatty acids, inflammation, and inflammatory diseases.
Am J Clin Nutr 83, 1505S–1519S | PubMedTsukada, H, Kakiuchi, T, Fukumoto, D, et al. (2000)
Docosahexaenoic acid (DHA) improves the age-related impairment of the coupling mechanism between
neuronal activation and functional cerebral blood flow response: a PET study in conscious monkeys.
Brain Res 862, 180–186Fontani, G, Corradeschi, A, Felici, F, et al. (2005)
Cognitive and physiological effects of omega-3 polyunsaturated fatty acid supplementation in healthy subjects.
Eur J Clin Invest 35, 691–699McNamara, RK, Able, J, Jandacek, R, et al. (2010)
Docosahexaenoic acid supplementation increases prefrontal cortex activation during sustained attention
in healthy boys: a placebo-controlled, dose-ranging, functional magnetic resonance imaging study.
Am J Clin Nutr 91, 1060–1067Steinbrink, J, Villringer, A, Kempf, F, et al. (2006)
Illuminating the BOLD signal: combined fMRI-fNIRS studies.
Magn Reson Imaging 24, 495–505Villringer, A & Dirnagl, U (1995)
Coupling of brain activity and cerebral blood-flow – basis of functional neuroimaging.
Cerebrovasc Brain Metab Rev 7, 240–276 | PubMedHoshi, Y, Kobayashi, N & Tamura, M (2001)
Interpretation of near-infrared spectroscopy signals: a study with a newly developed perfused rat brain model.
J Appl Physiol 90, 1657–1662 | PubMedFallgatter, AJ & Strik, WK (1997)
Right frontal activation during the continuous performance test assessed with near-infrared spectroscopy
in healthy subjects.
Neurosci Lett 223, 89–92Fallgatter, AJ & Strik, WK (1998)
Frontal brain activation during the Wisconsin Card Sorting Test assessed with two-channel
near-infrared spectroscopy.
Eur Arch Psych Clin Neurosci 248, 245–249Schecklmann, M, Ehlis, AC, Plichta, MM, et al. (2008)
Functional near-infrared spectroscopy: a long-term reliable tool for measuring brain activity during verbal fluency.
Neuroimage 43, 147–155Jasdzewski, G, Strangman, G, Wagner, J, et al. (2003)
Differences in the hemodynamic response to event-related motor and visual paradigms as measured by
near-infrared spectroscopy.
Neuroimage 20, 479–488Kanamaru, Y, Kikukawa, A, Miyamoto, Y, et al. (2008)
Dimenhydrinate effect on cerebral oxygen status and salivary chromogranin-A during cognitive tasks.
Prog Neuropsychopharmacol Biol Psychiatry 32, 107–115Bonoczk, P, Panczel, G & Nagy, Z (2002)
Vinpocetine increases cerebral blood flow and oxygenation in stroke patients:
a near infrared spectroscopy and transcranial Doppler study.
Eur J Ultrasound 15, 85–91Kennedy, DO, Wightman, EL, Reay, JL, et al. (2010)
Effects of resveratrol on cerebral blood flow variables and cognitive performance in humans:
a double-blind, placebo-controlled, crossover investigation.
Am J Clin Nutr 91, 1590–1597Kennedy, DO & Haskell, CF (2011)
Cerebral blood flow and behavioural effects of caffeine in habitual and non-habitual consumers
of caffeine: a near infrared spectroscopy study.
Biol Psychol 86, 298–306Hashimoto, M, Hossain, MS, Yamasaki, H, et al. (1999)
Effects of eicosapentaenoic acid and docosahexaenoic acid on plasma membrane fluidity
of aortic endothelial cells.
Lipids 34, 1297–1304Katayama, Y, Katsumata, T, Muramatsu, H, et al. (1997)
Effect of long-term administration of ethyl eicosapentate (EPA-E) on local cerebral blood flow
and glucose utilization in stroke-prone spontaneously hypertensive rats (SHRSP).
Brain Res 761, 300–305Jackson, PA, Deary, M, Reay, JL, et al. (2011)
No effect of 12 weeks' supplementation with 1 g DHA-rich or EPA-rich fish oil on cognitive function
or mood in healthy young adults aged 18–35 years.
Br J Nutr (epublication ahead of print version) doi:10.1017/S000711451100403XChance, B, Leigh, JS, Miyake, H, et al. (1988)
Comparison of time-resolved and time-unresolved measurements of deoxyhemoglobin in brain.
Proc Natl Acad Sci U S A 85, 4971–4975Vanderhasselt, MA, De Raedt, R & Baeken, C (2009)
Dorsolateral prefrontal cortex and Stroop performance: tackling the lateralization.
Psychon Bull Rev 16, 609–612Fitzgerald, PB, Srithiran, A, Benitez, J, et al. (2008)
An fMRI study of prefrontal brain activation during multiple tasks in patients with
major depressive disorder.
Hum Brain Mapp 29, 490–501Toone, BK, Okocha, CI, Sivakumar, K, et al. (2000)
Changes in regional cerebral blood flow due to cognitive activation among patients with schizophrenia.
Br J Psychiatry 177, 222–228Kennedy, DO, Veasey, R, Watson, A, et al. (2010)
Effects of high-dose B vitamin complex with vitamin C and minerals on subjective mood and performance
in healthy males.
Psychopharmacology 211, 55–68Hoshi, Y (2007)
Functional near-infrared spectroscopy: current status and future prospects.
J Biomed Opt 12, 062106Keppel, G (1991)
Design and Analysis.
Upper Saddle River, NJ: Prentice HallSublette, ME, Milak, MS, Hibbeln, JR, et al. (2009)
Plasma polyunsaturated fatty acids and regional cerebral glucose metabolism in major depression.
Prostaglandins Leukot Essent Fatty Acids 80, 57–64Huppert, TJ, Hoge, RD, Diamond, SG, et al. (2006)
A temporal comparison of BOLD, ASL, and NIRS hemodynamic responses to motor stimuli in adult humans.
Neuroimage 29, 368–382Kirby, A, Woodward, A, Jackson, S, et al. (2010)
A double-blind, placebo-controlled study investigating the effects of omega-3 supplementation
in children aged 8–10 years from a mainstream school population.
Res Dev Disabil 31, 718–730Osendarp, SJM, Baghurst, KI, Bryan, J, et al. (2007)
Effect of a 12-mo micronutrient intervention on learning and memory in well-nourished and marginally
nourished school-aged children: 2 parallel, randomized, placebo-controlled studies in Australia and Indonesia.
Am J Clin Nutr 86, 1082–1093
Return to OMEGA-3 FATTY ACIDS
Since 11-03-2019
| Home Page | Visit Our Sponsors | Become a Sponsor |
Please read our DISCLAIMER |